Biological Psychology 94 (2013) 592–600
DOI: 10.1016/j.biopsycho.2013.05.022
Authors
Péter Simora,∗ , Klára Horváthb,c , Péter P. Ujmab , Ferenc Gombosd , Róbert Bódizsb
Affiliations
a Department of Cognitive Sciences, Budapest University of Technology and Economics, Egry József u.1. Tépület/V, H-1111 Budapest, Hungary
b Institute of Behavioural Sciences, Semmelweis University, Nagyvárad tér 4, H-1089 Budapest, Hungary
c Department of Experimental Psychology, University of Oxford, South Parks Road, Oxford OX1 3UD, UK
d Department of General Psychology, Pázmány Péter Catholic University, Egyetem u. 1., 2087 Piliscsaba, Hungary
∗ Corresponding author at: H-1111 Egry József u.1. Tépület/V.em., Budapest, Hungary. Tel.: +36 1 463 1273; fax: +36 1 463 1072. E-mail addresses: psimor@cogsci.bme.hu, petersimor@gmail.com (P. Simor).
Keywords: Sleep Nightmare, Alpha oscillations, Power spectral analysis, REM, parasomnia
Abstract
Although a growing body of research indicates that frequent nightmares are related to impaired sleep regulation, the pathophysiology of nightmare disorder is far from being fully understood. We examined the relative spectral power values for NREM and REM sleep separately in 19 individuals with nightmare disorder and 21 healthy controls, based on polysomnographic recordings of the second nights’ laboratory sleep. Nightmare subjects compared to controls exhibited increased relative high alpha (10–14.5 Hz) and fronto-central increases in high delta (3–4 Hz) power during REM sleep, and a trend of increased fronto-central low alpha (7.75–9 Hz) power in NREM sleep. These differences were independent of the confounding effects of waking emotional distress. High REM alpha and low NREM alpha powers were strongly related in nightmare but not in control subjects. The topographical distribution and spectral components of REM alpha activity suggest that nightmare disordered subjects are characterized by wakelike electroencephalographic features during REM sleep.
Introduction
Nightmares are intense and highly unpleasant mental experiences that occur usually – but not exclusively – during late-night Rapid Eye Movement (REM) sleep and often provoke abrupt awakenings (ICSD-II, 2005). Nightmares affect approximately 4% of the adult population on a weekly basis (Spoormaker, Schredl, & van den Bout, 2006). Even though nightmares are often co-morbid with a wide variety of mental complaints (Levin & Nielsen, 2007), research indicates that frequent nightmares are more appropriate to be conceptualized as a specific sleep disorder that are independent in its origins from waking psychopathological symptoms (Coolidge, Segal, Coolidge, Spinath, & Gottschling, 2010; Lancee, Spoormaker, & van den, 2010; Spoormaker et al., 2006). Frequent nightmares are related to impaired subjective sleep quality in different agegroups and populations (Li et al., 2011; Li, Zhang, Li, & Wing, 2010; Schredl, 2010), and negatively toned dreams are more frequent among subjects with different sleep disorders (Schredl, 2009a,b; Schredl, Schafer, Weber, & Heuser, 1998) in whom nightmares seem to increase the severity of sleep complaints (Krakow, 2006; Schredl, 2009a,b). In consistence with questionnaire-based findings, early polysomnographic studies reported altered sleep architecture and sleep fragmentation in subjects with frequent nightmares (Fisher, Byrne, Edwards, & Kahn, 1970; Newell, Padamadan, & Drake, 1992). In addition, a recent study that also controlled for the confounding effects of co-morbid waking symptoms of depression and anxiety found decreased sleep efficiency, reduced slow wave sleep (SWS) and increased nocturnal awakenings in a group of young nightmare sufferers (Simor, Horváth, Gombos, Takács, & Bódizs, 2012). Others found enhanced periodic leg movements in nightmare sufferers with and without post-traumatic stress disorder suggesting that increased arousal and accompanying motor activation characterize the pathophysiology of nightmare disorder (Germain & Nielsen, 2003). Enhanced arousal during sleep was also evidenced by increased sympathetic (cardiac) activation in a group of nightmare subjects after a REM deprivation procedure (Nielsen et al., 2010). A more recent study found altered sleep microstructure in nightmare sufferers during Non-REM (NREM) sleep (Simor, Bódizs, Horváth, & Ferri, 2013) revealed by the Cyclic Alternating Pattern (CAP) analysis that quantifies and categorizes electroencephalographic (EEG) oscillations corresponding to recurrent activation events and transient states of unstable sleep depth (Terzano et al., 1985). More specifically, nightmare subjects in comparison to controls exhibited increased arousal responses comprised of desynchronized, high or mixed frequency activities and a reduced amount of synchronized, low frequency oscillations during spontaneous recurrent events of instability in NREM sleep and these differences were independent of the effects of anxious and depressive symptoms, indexed by psychometric tests. Desynchronized arousal responses that are generated usually at posterior sites and incorporate alpha (8–13 Hz) and beta (13–30 Hz) frequency bands shift the cortex toward a more alert brain state, while the generally antero-posterior propagation of slow (0.25–2 Hz) synchronized oscillations reflects the “effort” of the cortex to preserve sleep depth by reinforcing the thalamic-basal forebrain gate against arousing impulses (Parrino, Ferri, Bruni, & Terzano, 2012). Therefore, these findings indicate that nightmare disorder is characterized by inefficient sleep regulation and increased arousal responses that reduce the threshold for awakening. Interestingly, although nightmare disorder is considered to be a REM parasomnia (ICSD-II, 2005), anomalies in sleep continuity have been reported mainly during NREM sleep (Simor et al., 2013, 2012). Nevertheless, it is feasible that the above studies based on the visual scoring of sleep EEG could not capture the subtle alterations in the structure of neural oscillations during REM sleep. Power spectral analysis provides a fine-grained and sensitive examination of the electrophysiological oscillations during sleep, which seems to be an efficient tool to detect sleep alterations in different pathological conditions (Armitage, 1995; De la Fuente, Tugendhaft, & Mavroudakis, 1998; Feige, Scaal, Hornyak, Gann, & Riemann, 2007; Krystal, Edinger, Wohlgemuth, & Marsh, 2002; Lindberg et al., 2003; Moritz et al., 2002; Philipsen et al., 2005; Poulin, Stip, & Godbout, 2008). To the best of our knowledge, no previous studies have investigated the electrophysiological features of a whole night sleep in nightmare disorder. Therefore, our aim was to describe the EEG spectral profile of NREM and REM sleep in a group of nightmare subjects in comparison with that of controls.
Materials and methods
Participants
Participants (all native Hungarians) were selected from a large pool of undergraduate students from the Budapest University of Technology and Economics and Semmelweis University. Nightmare (NMs) and control subjects (CTLs) were enrolled after a stringent selection procedure described previously in detail (Simor et al., 2012). In brief, the enrollment was based on subjects’ scores on three different dreaming-related questionnaires: the Dream Quality Questionnaire (DQQ) (Bódizs, Simor, Csóka, Bérdi, & Kopp, 2008), the Hungarian version of the Van Dream Anxiety Scale (VDAS-H) (Simor et al., 2009) and two seven-point Likert scales, one assessing the frequency of awakening nightmares, and the other assessing the frequency of bad dreams without awakenings (0 – almost never; 1 – once or twice a year; 2 – every 2–3 month; 3 – once in a month; 4 – twice a month; 5 – once a week; 6 – more than once a week). NMs were selected on the basis of the International Classification of Sleep Disorders, 2nd edition (2005) criteria and Levin and Nielsen’s (2007) model of disturbed dreaming, including disturbed dreamers without abrupt awakenings. Subjects reporting one or more nightmares with awakening and/or bad dreams without awakening per week in the retrospective questionnaires were assigned to the NMs group, while individuals having less than two nightmares and/or bad dreams during the previous year were assigned as CTLs. Those subjects who reported the onset of negative dream experiences in relation to a traumatic event or indicated that the content of their dreams were related to a prior trauma (such as physical attack, accident, sudden death of a close relative, etc.) were excluded from the study. 21 NMs and 23 CTLs took part in polysomnographic examination; however, two NMs left the experiment after the first (baseline) night and two of the CTLs’ recordings were considered too noisy for spectral analyses. Therefore, 19 NMs (10 males; Mage = 20.87 ± 1.57) and 21 CTLs (11 male; Mage = 21.57 ± 1.47) were included in the present study. There was no significant age difference between the two groups (U(38) = 146; Z = −1.48; p = 0.138). NMs scored higher on the Negative Dream Affect Scale of the DQQ (MNMs = 8.12; SDNMs = 1.91 vs. MCTLs = 4.03; SDCTLs = 1.77; t(38) = −7.25; p < 0.0001) and on the VDAS-H (MNMs = 20.58; SDNMs = 7.5 vs. MCTLs : 0.23 ± SDCTLs = 0.62; t(18.22) = −11.73; p < 0.0001; equal variances not assumed), indicating at least moderately severe dream disturbances (Bódizs, Sverteczki, & Mészáros, 2008; Simor et al., 2009). None of the subjects reported prior neurological, psychiatric or sleep disorders or prior history of any chronic disease. The study protocol was approved by the Ethical Committee of the Semmelweis University. The subjects received monetary compensation for their participation in the sleep laboratory investigations. Written informed consent was obtained.
Procedure
Polysomnographic recordings were performed in the sleep research laboratory of the Semmelweis University for two consecutive nights. (The first night served as the adaptation night.) Subjects were not allowed to drink alcohol or take drugs (except contraceptives) on the day and the previous day of the examination. They were asked to avoid napping and consuming caffeine in the afternoon of the sleep recordings. The timing of lights off was between 11.00 PM and 1.00 AM depending on each participant’s preferred bedtime. Morning awakenings were scheduled after 9 h of undisturbed sleep unless participants woke up earlier spontaneously. Five of the NMs reported negatively toned dreams in the laboratory. In the morning – in order to measure subjective sleep quality – subjects were asked to complete the Hungarian adaptation of the Groningen Sleep Quality Scale (GSQS) (Simor, Köteles, Bódizs, & Bárdos, 2009). The one-dimensional 14-item questionnaire measures the extent of subjective sleep fragmentation. In order to control for the confounding effects of waking emotional distress on sleep EEG the Hungarian versions of the Spielberger Trait Anxiety Inventory (STAI-T) (Spielberger, Gorsuch, & Lushene, 1970) and the short Beck Depression Inventory (BDI-H) (Rózsa, Szádoczky, & Füredi, 2001) were assessed. The STAI-T is a widely used self-report instrument that differentiates between the temporary condition of state anxiety and the longstanding quality of trait anxiety. We used the 20-item Hungarian version of the STAI-T to assess general levels of anxiety (Sipos, Sipos, & Spielberger, 1994). The 9-item BDI-H is a one-dimensional scale assessing different symptoms of depression including social withdrawal, indecision, sleep disturbance, fatigue, intense worry about bodily symptoms, loss of work performance, pessimism, lack of satisfaction and self accusation (Rózsa et al., 2001).
Polysomnography
On both nights, subjects were fitted with 19 EEG electrodes (Fp1, Fp2, F3, F4, Fz, F7, F8, C3, C4, Cz, P3, P4, Pz, T3, T4, T5, T6, O1, O2) according to the 10–20 electrode placement system (Jasper, 1958) as well as with two EOG electrodes (bipolar channel) monitoring vertical and horizontal eye-movements; EMG electrodes (bipolar channels) for the chin and for the anterior tibialis muscles, two ECG electrodes according to standard lead I; in addition to the thoracic and abdominal respiration sensors. Gold-coated Ag/AgCl EEG cup electrodes were fixed with EC2 Grass Electrode Cream (Grass Technologies, USA) and referred to the mathematically-linked mastoids. Impedances were kept below 8 k[1]. Signals were collected, prefiltered (0.33–1500 Hz, 40 dB/decade anti-aliasing hardware input filter), amplified and digitized with 4096 Hz/channel sampling rate (synchronous) with 12 bit resolution by using the 32 channel EEG/polysystem (Brain-Quick BQ 132S, Micromed, Italy). A further 40 dB/decade anti-aliasing digital filter was applied by digital signal processing which low-pass filtered the data at 450 Hz. Finally, the digitized and filtered EEG was undersampled at 1024 Hz.
Spectral analyses
Sleep stages and conventional parameters of sleep macrostructure were scored according to Rechtschaffen and Kales standardized criteria (Rechtschaffen & Kales, 1968) by two experienced sleep researchers who were blind to the group membership of the participants. Overlapping (50%), artifact-free four-second-epochs of all EEG derivations were Hanning-tapered and Fourier transformed by using the FFT (Fast Fourier Transformation) algorithm in order to calculate the average power spectral densities (V2 /0.25 Hz) for whole night NREM (stages 2–4) and REM sleep periods. Since the absolute power values may be biased due to differences in the thickness – and thus the conductivity – of the skull, leading to disproportionate discrepancies between males and females (Carrier, Land, Buysse, Kupfer, & Monk, 2001), we applied the relative spectral power values. Relative spectral power values were obtained for each frequency bin (width: 0.25 Hz) by dividing the absolute power of the given frequency bin with the total spectral power (the sum of the absolute power of the whole range of analysis between 0.75 Hz and 48.25 Hz). The relative power values reflect the relative contribution of a given frequency range to the total spectrum. Relative spectral power values were log-transformed by using a 10 base logarithm in order to normalize their distribution before performing statistical analyses.
Statistical analyses
Statistical analyses were carried out with the Statistical Package for the Social Sciences version 19.0 (SPSS, IBM) and MATLAB (version 7.10.0.499, R2010a, The MathWorks, Inc., Natick, MA). Group differences of mean scores regarding the psychometric tests were compared with independent samples t-tests. If the psychometric parameters were not normally distributed, non-parametric (Mann–Whitney) tests were applied. Group comparisons of sleep architecture variables and relative spectral power values in each electrode and frequency bin were performed using multiple univariate analysis of covariance (ANCOVA). Relative spectral power values served as dependent variables, group membership was a fixed factor, and in order to control for the confounding effects of waking symptoms of anxiety and depressive states, STAI-T and BDI-H scores were used as covariates in the model. The sleep architecture variables that were not normally distributed were transformed to a natural logarithmic scale for statistical analysis. The level of significance was set at p < 0.05. Multiple comparisons inflate Type 1 error; however, statistical corrections ˇ correction) are too conservative for EEG data (such as the Bonferroni or the Sidák because they assume that the individual tests are independent from each other, which is not the case for EEG relative spectral power values. In order to address the issue of multiple comparisons we used the procedure of descriptive data analysis, delineating so called Rüger’s areas (Abt, 1987; Duffy et al., 1990). The procedure is designed to handle data with strongly intercorrelated neighboring data points. Rüger’s areas are defined as sets of conventionally significant (p < 0.05) results which are accepted or rejected as significant as a whole, instead of individual results of statistical tests. We took the results of our statistical tests as a two-dimensional matrix (with electrodes being one dimension, and frequency bins the other) and defined Rüger’s areas along the dimension of frequency bins. A Rüger’s area started when there was a significant result on any electrode in that frequency bin, continued into higher frequencies, and ended wherever there were no significant results on any electrode.
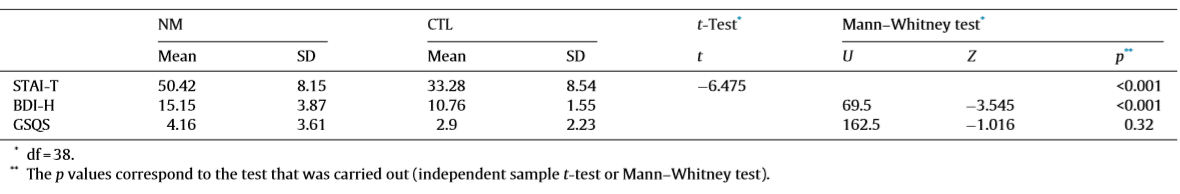
Table 1 Descriptive statistics and group comparisons of the psychometric variables
After defining these areas of significance, the number of significant results within the area was calculated, and it was investigated whether at least half of these results were significant at least at 1/2 of the conventional p = 0.05 significance level (that is, whether they were below 0.025) and at least one-third of them were significant at least at 1/3 of the conventional p = 0.05 significance level (that is, whether they were below 0.0167). If both of these conditions were fulfilled, the area as a whole was considered significant. With this method, a single significant statistical test with p < 0.0167 theoretically counts as a significant Rüger’s area. However, we did not consider single-bin results as an area. This conservative method addresses the issue of multiple comparisons since the probability that all these requirements will be fulfilled by chance is low.
Results
Psychometric tests and sleep macrostructure
The sleep macrostructure of an overlapping sample has already been reported in details elsewhere (Simor et al., 2012). However, as the sample of the present study was slightly different due to the exclusion of recordings that were too noisy for spectral analysis and due to the exclusion of recordings that were too noisy for spectral analysis and due to the broadening of the sample, we briefly describe the results. To analyze the group differences in terms of psychometric variables independent sample t-tests and Mann–Whitney tests (if the assumption of normality was violated) were conducted. To investigate the group main effect on sleep architecture, analysis of covariance was performed with STAI-T and BDI-H as covariates. Table 1 summarizes the descriptive statistics and the group comparisons of the psychometric variables, where NMs scored significantly higher both on the anxiety (STAI-T) and the depression (BDI-H) scales. In Table 2, the group differences regarding the sleep architecture can be observed. Even though no difference was found in subjective sleep quality of the two groups (GSQS), the objective sleep variables indicated impaired sleep architecture in NMs who had lower sleep efficiency and longer wake after sleep onset (WASO). Further, NMs were characterized by longer percentage of S1 sleep and lower proportion of SWS relative to the total sleep time.
Relative spectral power of NREM and REM sleepEEG
To analyze group differences in the EEG relative spectrum, analysis of covariance was carried out with relative power spectral density at the frequency bins as dependent variables and STAI-T and BDI-H as covariates. The differences between the relative spectra of NMs and CTLs can be observed in Fig. 1, where the data of the central midline (Cz) derivation are shown. Fig. 2A shows the proportion of the adjusted means of the two groups by electrode and by frequency band. Fig. 2B shows the p-values (only p < 0.15 are displayed) corresponding to the group main effects of the ANCOVA model. Significant Rüger’s areas surviving the statistical correction by descriptive data analysis are indicated in the figure.
Clear differences can be observed both in NREM and REM sleep. Regarding NREM sleep, NMs showed significantly lower 1–1.25 Hz activity in various recording sites (F8, C3, Cz, T4, P3, Pz, O1, O2), whereas they had a higher 4–4.75 Hz activity on the Cz and the Pz electrodes compared to CTLs. Furthermore, increased
Table 2 Descriptive statistics and group comparisons of the sleep architecture variables.
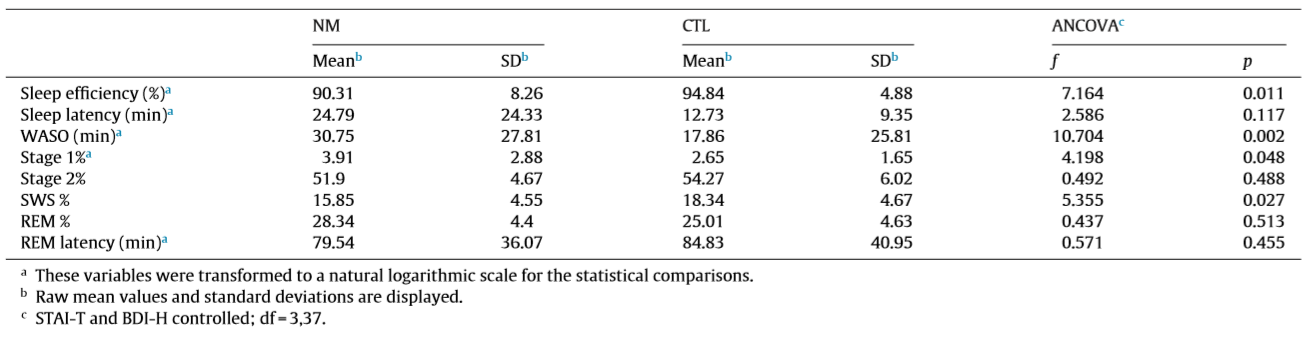
Fig. 1. Relative power spectra in NMs and CTLs. Group means of relative power spectra of NREM and REM sleep EEG at derivation Cz. The y axis shows the adjusted means (STAI = 41.43; BDI = 12.85) of the logarithmized raw data after back-transformation (10x ) for visualization. Significant differences before and after descriptive data analysis can be observed in Fig. 2. In sum, we found a tendency in the difference between the two groups regarding the NREM sleep EEG 7.75–9 Hz activity and a significant difference regarding the REM sleep EEG 3–4 and 10–14.5 Hz activity.
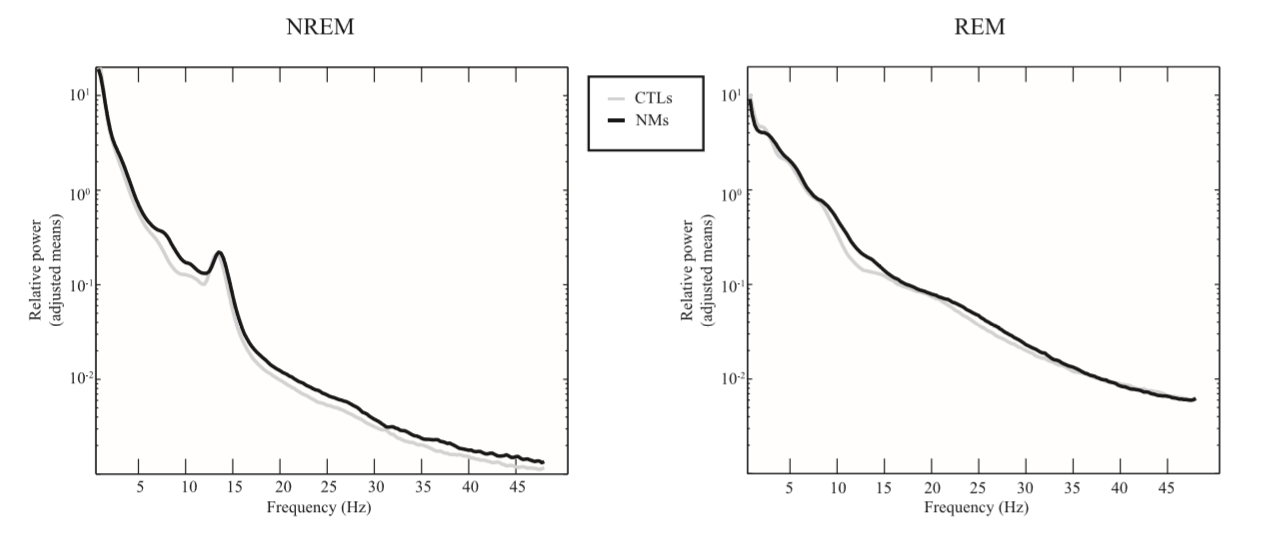
Fig. 2. Statistical difference between relative power spectra in NMs and CTLs. The proportion of NREM and REM sleep EEG relative power spectra in the two groups by electrodes (A) and the corresponding p-values of the ANCOVA group main effect (B). The proportion of adjusted means (STAI = 41.43; BDI = 12.85) are shown in A (the logarithmized raw data were back-transformed (10x ) for visualization). The horizontal lines below the graphs (gray line – tendency, black line – significant) and the vertical dashed lines indicate the frequency bins which were significant after the correction for multiple comparisons.
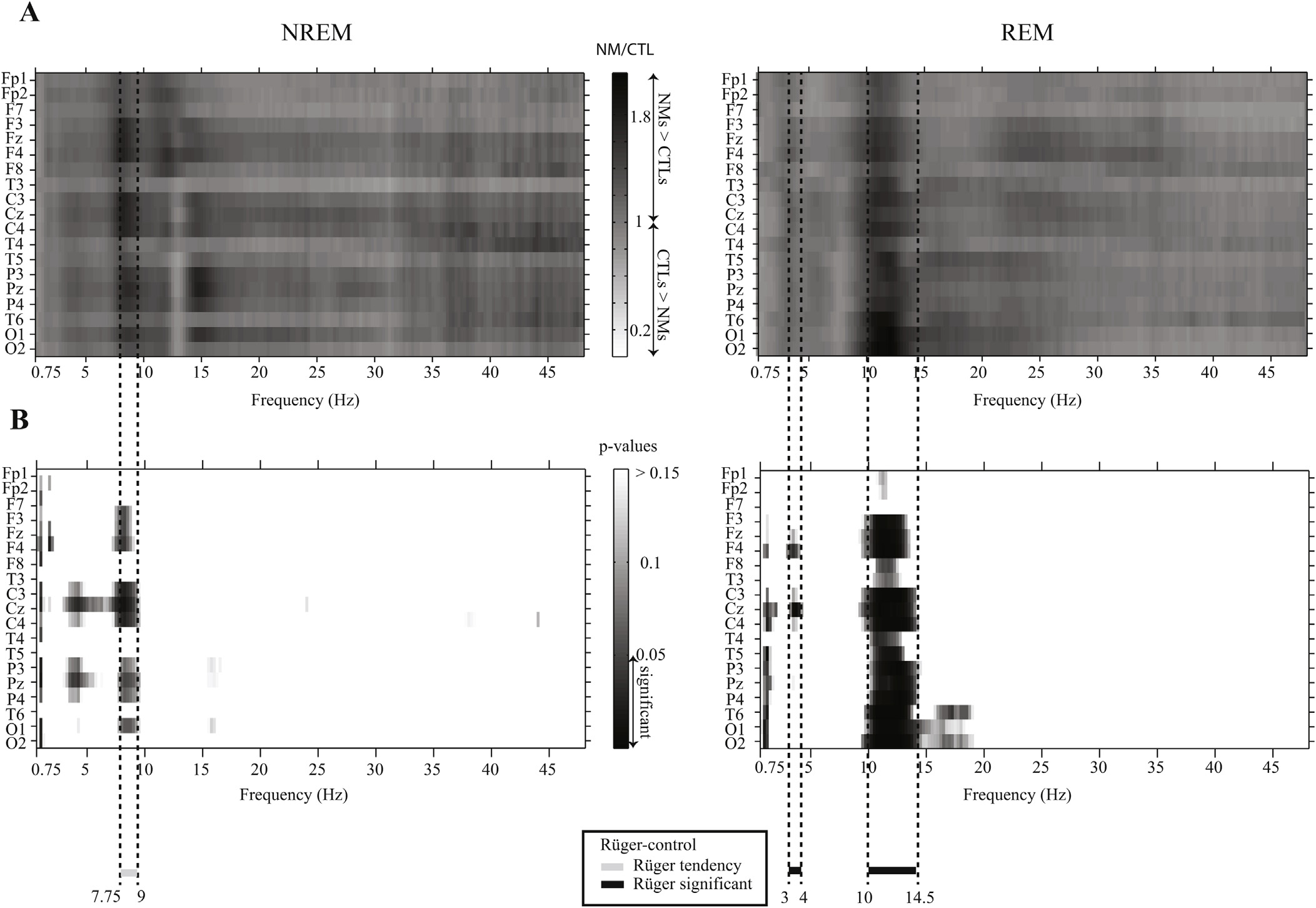
Fig. 3. Topographic distribution of REM high alpha activity. The topographic distribution of the REM sleep EEG 10–14.5 Hz activity in NMs and CTLs, as well as the difference between the two groups. Means of the raw relative power spectrum are displayed.
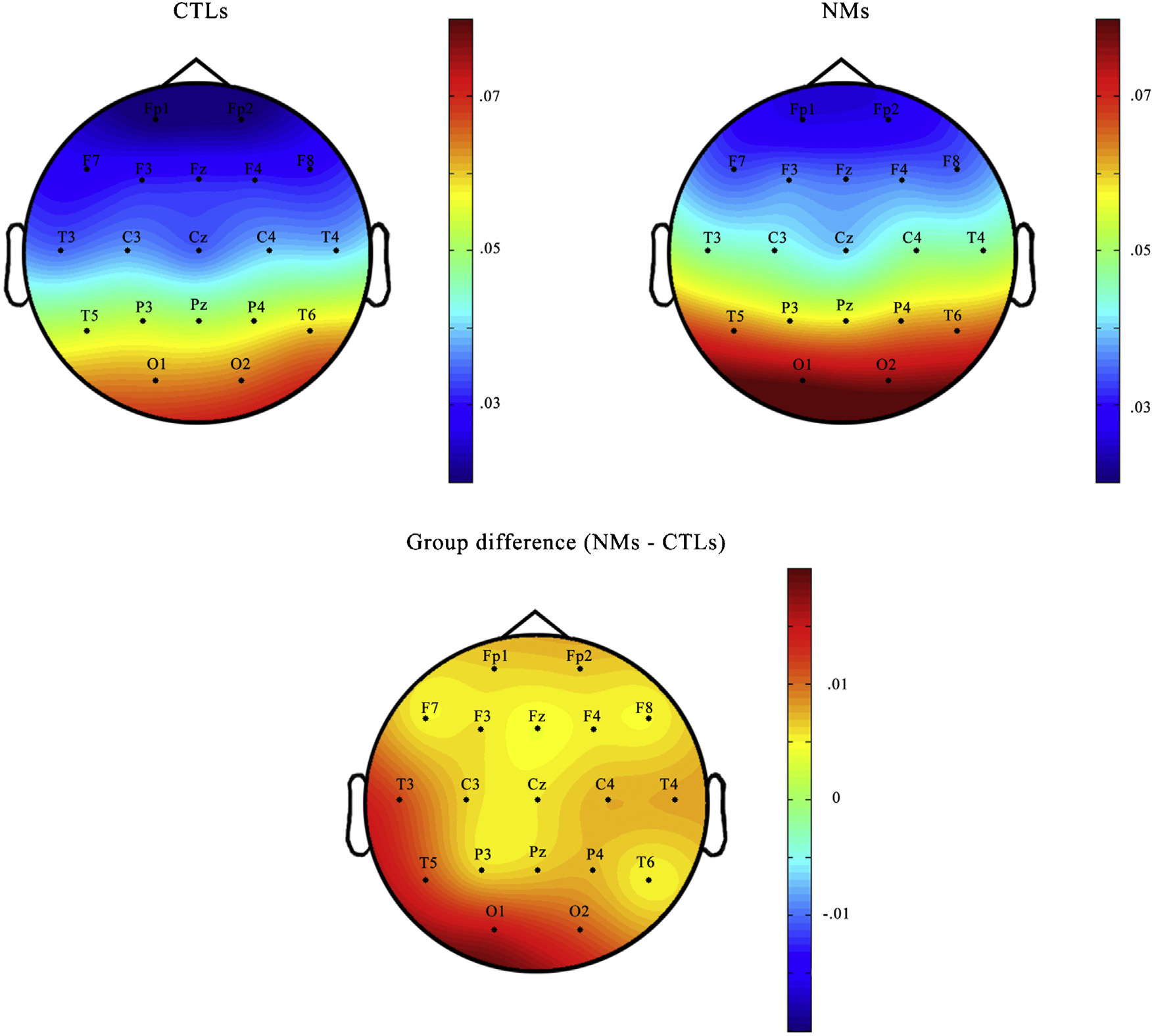
fronto-central (Fz, F4, C3, Cz, C4) 7.75–9 Hz activity was found in NMs. After the correction for multiple comparisons the differences in 1–1.25 Hz, 4–4.75 Hz frequency range were no more significant. Differences regarding the 7.75–9 Hz (low alpha range) showed a trend (one-quarter instead of one-half of the bins (20) were significant at the p < 0.025 level) (see Fig. 2B on the left). Regarding REM sleep, NMs were characterized by a significantly higher 10–14.5 Hz EEG activity almost all over the scalp (F3, Fz, F4, F8, C3, Cz, C4, T4, T5, P3, Pz, P4, T6, O1, O2) in comparison with CTLs, in addition to a fronto-central (F4, Cz) increase in the 3–4 Hz activity range. Differences in both frequency ranges remained significant after correction for multiple comparisons. To investigate further the topographic distribution of the 10–14.5 Hz activity in REM sleep that traditionally belongs to the high alpha range (Cantero, Atienza, & Salas, 2002), in Fig. 3 we plotted the relative power spectral density of this range for NMs and CTLs, as well as the difference between the two groups. The 10–14.5 Hz high alpha activity was prominent at the posterior sites, peaking at the occipital (O1, O2) derivations. According to Fig. 3, it is clear that the most significant differences between the groups emerged at the left temporo-occipital area. To sum up, a slight increase in fronto-central high delta as well as a prominent and expanded high alpha increase were found in the REM sleep of NMs along with a tendency of more concentrated NREM alterations regarding the low alpha frequency components.
Relationship between NREM low and REM high alpha EEG in NMs and CTLs
In order to examine if the tendency-like increase in EEG low alpha activity in the NREM phase of sleep and the significant increase in high alpha EEG activity in REM sleep that were observed in NMs were related to each other, we performed a post hoc correlation analysis within the two groups separately. Relative spectral power values were averaged over the derivations and Pearson product–moment correlations were computed between NREM 7.75–9 Hz and REM 10–14.5 Hz activity. In the NMs group, a significant and strong correlation (r(19) = 0.78; p = 0.0001) emerged, whereas in CTLs no significant correlation was found (r(21) = 0.22; p = 0.36) between the low alpha frequency range of NREM and the high alpha frequency range of REM sleep. In order to examine the relationship between NREM low/REM high alpha power with subjective sleep quality we performed Pearson product–moment correlations between these spectral measures and subjective sleep quality indexed by the GSQS scores. The spectral measures did not show significant correlations with
Table 3. Group and Gender effects on averaged relative spectral power values for different frequency domains. Frequency domain (relative spectral power)
the subjective sleep quality neither across nor within the two groups.
Temporal dynamics in NREM and REM relative spectral power
In order to examine the temporal dynamics of altered NREM and REM sleep in NMs, we calculated the relative spectral power for the first three NREM/REM cycles separately. After the control for multiple comparisons significant differences emerged in the first and the second NREM period. NMs showed reduced power in the 1–1.25 Hz frequency range in several recording sites (F7, C3, Cz, T5, P3, O1, O2) in the first, and increased power in the low alpha band, between 7.25 Hz and 9.25 Hz in the second NREM period in fronto-central derivations (Fp1, Fp2, F7, F3, Fz, F4, C3, Cz, C4). Regarding REM sleep significant group differences were found only in the high alpha range in all the REM periods. Nevertheless, the significant frequency ranges showed slight variations among the first three REM cycles (8.75–13.75 Hz, 9–16 Hz and 10–14.25 Hz in the first, second and third REM period, respectively).
Gender differences
We examined the influence of gender as well as the Group × Gender interactions on relative spectral power data. In order to reduce the number of parameters, we averaged relative spectral power values over scalp derivations and summed up frequency bins to generate six frequency band windows for NREM and REM sleep separately. Frequency domains included delta (0.75–4 Hz), theta (4.25–7.5 Hz), alpha (7.75–10.75 Hz), sigma (11–15 Hz), beta (15.25–31 Hz) and gamma (31.25–48.25 Hz) for NREM, and delta (0.75–4 Hz), theta (4.25–7.5 Hz), low alpha (7.75–9.75 Hz), high alpha (10–15 Hz), beta (15.25–31 Hz) and gamma (31.25–48.25 Hz) relative spectral power values for REM sleep. Multiple univariate ANCOVAs were performed with the above frequency bands for NREM and REM sleep as dependent, Group and Gender as independent and STAI-T and BDI-H scores as covariate variables in the models. To address the issue of multiple comparisons, we corrected the ␣-level with the number of frequency bands (Bonferroni correction). This way, after the correction for multiple comparisons the error rate was set to p = 0.05/#measures, or 0.05/12 = 0.0042. Detailed results are presented in Table 3. Regarding NREM sleep the effect of Gender was significant for the theta and alpha and delta frequency bands. Females exhibited higher values in the theta (Estimated Marginal Mean (EMM)females : 0.47; SE: 0.02 vs. EMMmales : 0.39; SE: 0.02) and alpha (EMMfemales : 0.15; SE: 0.01 vs. EMMmales : 0.11; SE: 0.01) and slightly lower values in delta (EMMfemales : 5.45; SE: 0.04 vs. EMMmales : 5.6; SE: 0.05) power in comparison with male subjects. In coherence with the bin-wise analysis, the effect of Group showed a trend in the alpha range, and a significant effect emerged for the interaction of Group and Gender, the latter due to increased alpha power in female NMs (Raw Mean: 0.18; SD: 0.08) in contrast to male NMs (Raw Mean: 0.08; SD: 0.03). Nevertheless, none of these differences survived the Bonferroni correction for multiple comparisons.
In REM sleep the effect of Group (EMMNMs : 0.45; SE: 0.03 vs. EMMCTLs : 0.29; SE: 0.03) and Gender (EMMfemales : 0.42; SE: 0.02 vs. EMMmales : 0.33; SD: 0.03) were significant for the high alpha frequency band, but only the effect of Group survived the correction for multiple comparisons. The interaction of Group and Gender was not significant for any of the examined frequency domains.
Discussion
To the best of our knowledge, the present study is the first to describe alterations in EEG spectral power in nightmare disorder. The most prominent finding is that subjects with frequent nightmares exhibited increased relative spectral power in the high alpha range (10–14.5 Hz) compared to healthy controls during REM sleep. In addition, increased REM high delta (3–4 Hz) activity and a tendency of increased NREM low alpha (7.75–9 Hz) activity were observed in NMs in contrast to CTLs. Separate analyses of the first three NREM/REM cycles revealed time dependent effects for the low alpha power in NREM sleep, showing significantly increased power in NMs only in the second NREM period. In contrast, high alpha power was increased in all the first three REM periods in NMs. Since these differences were statistically independent of the confounding effects of waking emotional distress indicative of subclinical psychopathological states, altered sleep microstructure seems to be an inherent feature of the neurophysiology of nightmare disorder.
We suggest that increased high alpha power in the REM periods of NMs is a wake-like feature during REM sleep which might contribute to the pathophysiology of nightmare disorder. Alpha oscillations during REM sleep might reflect relatively short periods of sleep instability (micro-arousals) that facilitate the connection between the sleeping brain and the external environment (Cantero, Atienza, & Salas, 2000; Halász, 1998; Halász, Terzano, Parrino, & Bódizs, 2004). Alpha oscillations were shown to be modulated differently in the different states of alertness (Cantero et al., 2002): while higher alpha components are dominant during wakefulness, REM sleep is characterized by the preponderance of slower (7.5–10.5 Hz) alpha oscillations in healthy subjects (Cantero, Atienza, Gómez, & Salas, 1999). Moreover, there are clear differences in the topographical distribution of the alpha activity between wakefulness and REM sleep. Posterior dominance of alpha power is characteristic of relaxed wakefulness, whereas in REM the distribution seems to be more homogeneous (Cantero et al., 1999, 2002). Therefore, the increased high alpha activity peaking at posterior locations in the REM periods of NMs may reflect a “hybrid state” with the occurrence of wake-type alpha oscillations during REM sleep.
In addition to the increased high alpha activity in REM sleep, the alpha activity in NREM sleep was also higher in NMs, but at lower frequencies (7.75–9 Hz), which may indicate increased alertness during NREM. A growing body of research suggests that alpha activity reflects an “internal” preparatory state that facilitates alertness and readiness for sensorimotor and cognitive processing (Linkenkaer-Hansen, Nikulin, Palva, Ilmoniemi, & Palva, 2004; Palva, Linkenkaer-Hansen, Näätänen, & Palva, 2005; Palva & Palva, 2007; Sadaghiani et al., 2010; Schürmann & Bas¸ar, 2001). A recent study by McKinney and colleagues (McKinney, Dang-Vu, Buxton, Solet, & Ellenbogen, 2011) extended this concept to the sleeping state by showing that the continuous variation of alpha power is a sensitive marker of sleep fragility and environmental awareness during NREM sleep. In light of these findings, we suggest that the slightly enhanced alpha power in NMs sleep reflects disturbed sleep regulation and increased alertness toward environmental stimuli during NREM sleep, especially in the first part (second sleep cycle) of the night. Furthermore, we showed that the increase in different alpha components during NREM and REM sleep were related, but only within the NMs group, suggesting that enhanced alpha power is a peculiar characteristic of the sleep pathophysiology of nightmare disorder; however, its frequency range seems to be modulated in a sleep state-dependent manner. In addition to group differences, gender also had an effect on spectral power measures. In coherence with previous findings on gender differences (Carrier et al., 2001), females exhibited higher values in NREM theta and alpha power. Nevertheless, a trend of Group and Gender interaction emerged for increased NREM alpha power. Females were shown to report more nightmares and bad dreams in both the general and clinical population, which might be related to trait-like differences in emotional reactivity (Levin & Nielsen, 2007). Although in our principal analyses we treated male and female NMs as a whole group, our findings revealed gender differences especially in NREM sleep spectral power. This suggests that proneness to nightmarish experiences may stem from different pathophysiological background in men and women.
Wake-like features in neural oscillations might have important consequences on mental experiences during sleep. Enhanced processing of the environment, including external (e.g. noise) as well as internal (e.g. proprioceptive) information may lead to insomniac complaints – like in sleep-state misperception (Riemann et al., 2010), but perhaps also to the intensification of dream experiences. We propose that the appearance of wake-type alpha oscillations during sleep – especially during the activated cortical state of REM sleep – might promote the intensification of sensorial, emotional and cognitive processes shaping the oneiric experience and result in perceptually vivid, realistic and emotionally absorbing dream images. Moreover, transient states of wake-like functioning may strengthen the memory traces for these dream experiences. It is worth noting that high, but not low alpha EEG oscillations involving the thalamo-cortical feed-back loops were hypothesized to reflect search and retrieval in semantic long-term memory during wakefulness (Klimesch, 1999). Thus, the significantly increased REM sleep high alpha EEG activity in NMs could reflect a permanent REM sleep-related intensification of cognitive activity possibly resulting in frequent nightmare experiences. These assumptions are in line with earlier studies showing the association between alpha activity in REM sleep and vivid dream experiences (Robert, Harry, Tyson, Melodie, & Daniel, 1982; Tyson, Ogilvie, & Hunt, 1984). Furthermore, nightmares seem to belong to a wide domain of unusual dream experiences like sleep paralysis, vivid dreaming, lucid dreaming or terrifying hypnagogic hallucinations, which usually occur during sleep–wake transitions (Nielsen & Zadra, 2011). Nevertheless, the question how the valence of these vivid dream experiences in NMs shifts toward the negative domain remains unanswered.
Our findings showing increased low NREM alpha/high REM alpha power in NMs might indicate that altered relative spectral power in NMs resembles sleep patterns found in insomnia; however, in case of insomniac subjects, conclusive findings emerged only during NREM sleep, showing decreased delta, and increased alpha, beta and sigma power (Krystal et al., 2002; Spiegelhalder et al., 2012), while there has been no convincing evidence on REM sleep alterations (Riemann et al., 2010). Moreover, in our sample neither NREM low alpha power, nor REM high alpha power correlated with subjective sleep quality scores. These results make unlikely that increased low NREM and high REM alpha power in NMs are related to concomitant symptoms of insomnia. Nevertheless, since nightmares and insomniac symptoms were reported to be associated in several studies (Li et al., 2011, 2010; Schredl, 2009a,b) proneness to insomniac complaints or subclinical insomniac symptoms might also play a role in shaping NMs’ sleep.
The neural mechanism of increased alpha power during sleep is far from being fully understood; however, research suggests that alpha activity is related to an alertness network comprising the dorsal anterior cingulate cortex, anterior insula, and thalamus relaying sensory stimuli to cortical processing (Sadaghiani et al., 2010). Furthermore, these fluctuations between sleep and wakeful states might be modulated by ascending monoaminergic pathways that increase the responsiveness of cortical and thalamic neurons to sensory information (Steriade, McCormick, & Sejnowski, 1993). The effect of different monoaminergic neurotransmitters on sleep EEG and dream intensity awaits further investigations; however, clinical research showed that an ␣-1 adrenergic antagonist agent (prazosin) reduced nightmares in patients with post-traumatic stress disorder (Dierks, Jordan, & Sheehan, 2007; Peskind, Bonner, Hoff, & Raskind, 2003; Raskind et al., 2007, 2003; Taylor & Raskind, 2002), while selective serotonin and norepinephrine reuptake inhibitors seem to increase the vividness and nightmarish quality of dreaming (Tribl, Wetter, & Schredl, 2012). NMs also exhibited increased power in 3–4 Hz high delta activity during REM sleep. Increased power in the high delta range during REM sleep is not easy to interpret; however, it is possible that it is related to an increased homeostatic sleep pressure (Marzano, Ferrara, Curcio, & Gennaro, 2010), the demand of the cortex to exhibit the thalamo-cortically generated “bursting mode” of hyperpolarization and rebound sequences (Steriade & Llinás, 1988) that could not be completely expressed due to inefficient sleep regulation and reduced slow wave sleep (Simor et al., 2012). An alternative view of the increased REM sleep 3–4 Hz activity in NMs might be related to the findings of Germain and Nielsen (2001), who reported a strong relationship between sleep onset hypnagogic imagery and EEG delta power. Given the similarities between sleep onset and REM sleep (Bódizs, Sverteczki, et al., 2008), as well as our major finding showing the intrusion of alpha-wave associated wakefulness-like processes into the course of REM sleep this hypothesis seems to be viable. We should note however, that increments in delta power were only evident at two recording sites (F4, Cz), therefore this finding should be treated carefully.
In contrast to our previous results showing impaired sleep regulation by decreased slow wave sleep (Simor et al., 2012) and reduced number of delta bursts (CAP A1) during arousal events (Simor et al., 2013), relative spectral power analysis did not reveal significant decrease in slow oscillatory activity surviving the correction for multiple comparisons. However, the quantitative EEG analysis is based on artifact free epochs (since movement related artifacts are excluded from the analyses), while sleep staging and CAP analysis include these periods as well. Therefore it is possible that decreased slow oscillatory activity in NMs is principally apparent during transient events of arousal and is less evident during the “restored” background activity that spectral power measures analyze. Furthermore, due to the moderate presence of sweating artifacts in some of our subjects our analysis was restricted to the frequency range above 0.75 Hz, thus our study does not provide information about the slow oscillatory activity under this frequency bin which might be altered in NMs. Nevertheless, decreases in NREM slow delta (1–1.25 Hz) and increases in NREM low theta (4–4.75 Hz) – although were non-significant after the statistical correction – might similarly reflect impaired sleep regulation in NMs. Regarding the limitations of our study we should note, that since we did not wake up our subjects during the night in order to collect dream reports, we cannot make direct inferences about the association between sleep EEG oscillations and nightmare formation. Hence, the direct relationship between sleep–wake transitions and mental experiences needs to be investigated further. Since nightmares do not occur every night and reports of nightmares are scarce in the sleep laboratory environment, assessments with more nights or ambulatory (home) recordings would provide a more efficient way to examine the neurophysiological background of nightmarish experiences. Examining a relatively homogeneous sample may reduce the influence of different confounding factors, even though we should be careful with generalizing our data to other age groups and populations. Further studies with larger sample sizes involving older subjects might help to resolve these limitations. Although we statistically controlled the effects of waking psychopathological symptoms on sleep EEG patterns, more research is warranted in order to examine the influence of more severe, co-morbid psychopathological symptoms on spectral power measures. Moreover, the influence of anxious or other pathological mood states should be investigated in further studies comparing nightmare and non-nightmare subsamples with high and low waking emotional distress. In spite of these shortcomings, our study reports novel data regarding the pathophysiology of nightmare disorder and provides testable hypotheses for further investigations examining the relationship between neural oscillations and mental experiences during sleep.
Acknowledgements
This research was realized in the frames of TÁMOP 4.2.4. A/111-1-2012-0001 “National Excellence Program – Elaborating and operating an inland student and researcher personal support system”. The project was subsidized by the European Union and co-financed by the European Social Fund. The present research was also supported by the 2010 Research Grant of the BIAL Foundation (55/10) and the 2009 Research Grant Award of the Joint IASD/DreamScience Foundation.
References
Abt, K. (1987). Descriptive data analysis: A concept between confirmatory and exploratory data analysis. Methods of Information in Medicine, 26(2), 77–88.
American Academy of Sleep Medicine. (2005). The international classification of sleep disorders: Diagnostic and coding manual (2nd ed.). Westchester: American Academy of Sleep Medicine.
Armitage, R. (1995). Microarchitectural findings in sleep EEG in depression: Diagnostic implications. Biological Psychiatry, 37(2), 72–84.
Bódizs, R., Simor, P., Csóka, S., Bérdi, M., & Kopp, M. S. (2008). Dreaming and health promotion: A theoretical proposal and some epidemiological establishments. European Journal of Mental Health, 3(1), 35–62.
Bódizs, R., Sverteczki, M., & Mészáros, E. (2008). Wakefulness–sleep transition: Emerging electroencephalographic similarities with the rapid eye movement phase. Brain Research Bulletin, 76(1–2), 85–89.
Cantero, J. L., Atienza, M., Gómez, C., & Salas, R. M. (1999). Spectral structure and brain mapping of human alpha activities in different arousal states. Neuropsychobiology, 39(2), 110–116.
Cantero, J. L., Atienza, M., & Salas, R. M. (2000). Spectral features of EEG alpha activity in human REM sleep: Two variants with different functional roles? Sleep, 23(6), 746–750.
Cantero, J. L., Atienza, M., & Salas, R. M. (2002). Human alpha oscillations in wakefulness, drowsiness period, and REM sleep: Different electroencephalographic phenomena within the alpha band. Neurophysiologie Clinique/Clinical Neurophysiology, 32(1), 54–71.
Carrier, J., Land, S., Buysse, D. J., Kupfer, D. J., & Monk, T. H. (2001). The effects of age and gender on sleep EEG power spectral density in the middle years of life (ages 20–60 years old). Psychophysiology, 38(2), 232–242.
Coolidge, F. L., Segal, D. L., Coolidge, C. M., Spinath, F. M., & Gottschling, J. (2010). Do nightmares and generalized anxiety disorder in childhood and adolescence have a common genetic origin?? Behavior Genetics, 40(3), 349–356.
De la Fuente, J. M., Tugendhaft, P., & Mavroudakis, N. (1998). Electroencephalographic abnormalities in borderline personality disorder. Psychiatry Research, 77(2), 131–138.
Dierks, M. R., Jordan, J. K., & Sheehan, A. H. (2007). Prazosin treatment of nightmares related to posttraumatic stress disorder. The Annals of Pharmacotherapy, 41(6), 1013–1017.
Duffy, F. H., Jones, K., Bartels, P., Albert, M., McAnulty, G. B., & Als, H. (1990). Quantified neurophysiology with mapping: Statistical inference, exploratory and confirmatory data analysis. Brain Topography, 3(1), 3–12.
Feige, B., Scaal, S., Hornyak, M., Gann, H., & Riemann, D. (2007). Sleep electroencephalographic spectral power after withdrawal from alcohol in alcohol-dependent patients. Alcoholism, Clinical and Experimental Research, 31(1), 19–27.
Fisher, C., Byrne, J., Edwards, A., & Kahn, E. (1970). A psychophysiological study of nightmares. Journal of the American Psychoanalytic Association, 18, 747–782.
Germain, A., & Nielsen, T. A. (2001). EEG power associated with early sleep onset images differing in sensory content. Sleep Research Online, 4(3), 83–90.
Germain, A., & Nielsen, T. A. (2003). Sleep pathophysiology in posttraumatic stress disorder and idiopathic nightmare sufferers. Biological Psychiatry, 54(10), 1092–1098.
Halász, P. (1998). Hierarchy of micro-arousals and the microstructure of sleep. Neurophysiologie Clinique/Clinical Neurophysiology, 28(6), 461–475.
Halász, P., Terzano, M., Parrino, L., & Bódizs, R. (2004). The nature of arousal in sleep. Journal of Sleep Research, 13(1), 1–23.
Jasper, H. (1958). Report of the committee on methods of clinical examination in electroencephalography. Electroencephalography and Clinical Neurophysiology, 10, 370–375.
Klimesch, W. (1999). EEG alpha and theta oscillations reflect cognitive and memory performance: A review and analysis. Brain Research Reviews, 29(2), 169–195.
Krakow, B. (2006). Nightmare complaints in treatment-seeking patients in clinical sleep medicine settings: Diagnostic and treatment implications. Sleep, 29(10), 1313–1319.
Krystal, A. D., Edinger, J. D., Wohlgemuth, W. K., & Marsh, G. R. (2002). NREM sleep EEG frequency spectral correlates of sleep complaints in primary insomnia subtypes. Sleep, 25(6), 630–640.
Lancee, J., Spoormaker, V. I., & van den, B. (2010). Nightmare frequency is associated with subjective sleep quality but not with psychopathology. Sleep and Biological Rhythms, 8(3), 187–193.
Levin, R., & Nielsen, T. A. (2007). Disturbed dreaming, posttraumatic stress disorder, and affect distress: A review and neurocognitive model. Psychological Bulletin, 133(3), 482–528.
Li, S. X., Yu, M. W. M., Lam, S. P., Zhang, J., Li, A. M., Lai, K. Y. C., et al. (2011). Frequent nightmares in children: Familial aggregation and associations with parent-reported behavioral and mood problems. Sleep, 34(4), 487–493.
Li, S. X., Zhang, B., Li, A. M., & Wing, Y. K. (2010). Prevalence and correlates of frequent nightmares: A community-based 2-phase study. Sleep, 33(6), 774–780.
Lindberg, N., Tani, P., Appelberg, B., Naukkarinen, H., Rimón, R., Porkka-Heiskanen, T., et al. (2003). Human impulsive aggression: A sleep research perspective. Journal of Psychiatric Research, 37(4), 313–324.
Linkenkaer-Hansen, K., Nikulin, V. V., Palva, S., Ilmoniemi, R. J., & Palva, J. M. (2004). Prestimulus oscillations enhance psychophysical performance in humans. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience, 24(45), 10186–10190.
Marzano, C., Ferrara, M., Curcio, G., & Gennaro, L. D. (2010). The effects of sleep deprivation in humans: Topographical electroencephalogram changes in nonrapid eye movement (NREM) sleep versus REM sleep. Journal of Sleep Research, 19(2), 260–268.
McKinney, S. M., Dang-Vu, T. T., Buxton, O. M., Solet, J. M., & Ellenbogen, J. M. (2011). Covert waking brain activity reveals instantaneous sleep depth. PLOS ONE, 6(3), e17351.
Moritz, S., Birkner, C., Kloss, M., Jahn, H., Hand, I., Haasen, C., et al. (2002). Executive functioning in obsessive–compulsive disorder, unipolar depression, and schizophrenia. Archives of Clinical Neuropsychology, 17(5), 477–483.
Newell, S. A., Padamadan, H., & Drake, M. E., Jr. (1992). Neurophysiologic studies in nightmare sufferers. Clinical EEG (Electroencephalography), 23(4), 203–206.
Nielsen, T., Paquette, T., Solomonova, E., Lara-Carrasco, J., Colombo, R., & Lanfranchi, P. (2010). Changes in cardiac variability after REM sleep deprivation in recurrent nightmares. Sleep, 33(1), 113–122.
Nielsen, T., & Zadra, A. (2011). Idiopathic nightmares and dream disturbances associated with sleep–wake transitions. In M. Kryger, T. Roth, & W. C. Dement (Eds.), Principles and practice of sleep medicine (5th ed., pp. 1106–1115). New York: Elsevier.
Palva, S., Linkenkaer-Hansen, K., Näätänen, R., & Palva, J. M. (2005). Early neural correlates of conscious somatosensory perception. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience, 25(21), 5248–5258.
Palva, S., & Palva, J. M. (2007). New vistas for alpha-frequency band oscillations. Trends in Neurosciences, 30(4), 150–158.
Parrino, L., Ferri, R., Bruni, O., & Terzano, M. G. (2012). Cyclic alternating pattern (CAP): The marker of sleep instability. Sleep Medicine Reviews, 16(1), 27–45.
Peskind, E. R., Bonner, L. T., Hoff, D. J., & Raskind, M. A. (2003). Prazosin reduces trauma-related nightmares in older men with chronic posttraumatic stress disorder. Journal of Geriatric Psychiatry and Neurology, 16(3), 165–171.
Philipsen, A., Feige, B., Al-Shajlawi, A., Schmahl, C., Bohus, M., Richter, H., et al. (2005). Increased delta power and discrepancies in objective and subjective sleep measurements in borderline personality disorder. Journal of Psychiatric Research, 39(5), 489–498.
Poulin, J., Stip, E., & Godbout, R. (2008). REM sleep EEG spectral analysis in patients with first-episode schizophrenia. Journal of Psychiatric Research, 42(13), 1086–1093.
Raskind, M. A., Peskind, E. R., Hoff, D. J., Hart, K. L., Holmes, H. A., Warren, D., et al. (2007). A parallel group placebo controlled study of prazosin for trauma nightmares and sleep disturbance in combat veterans with post-traumatic stress disorder. Biological Psychiatry, 61(8), 928–934.
Raskind, M. A., Peskind, E. R., Kanter, E. D., Petrie, E. C., Radant, A., Thompson, C. E., et al. (2003). Reduction of nightmares and Other PTSD Symptoms in combat veterans by prazosin: A placebo-controlled study. American Journal of Psychiatry, 160(2), 371–373.
Rechtschaffen, A., & Kales, A. (1968). A manual of standardized terminology, technique and scoring system for sleep stages of human sleep. Los Angeles Brain Information Service. Brain Information Institute, UCLA.
Riemann, D., Spiegelhalder, K., Feige, B., Voderholzer, U., Berger, M., Perlis, M., et al. (2010). The hyperarousal model of insomnia: A review of the concept and its evidence. Sleep Medicine Reviews, 14(1), 19–31.
Robert, D. O., Harry, T., Tyson, P. D., Melodie, L. L., & Daniel, B. J. (1982). Lucid dreaming and alpha activity: A preliminary report. Perceptual and Motor Skills, 55(3), 795–808.
Rózsa, S., Szádoczky, E., & Füredi, J. (2001). A Beck Depresszió Kérdőív rövidített változatának jellemzői a hazai mintán. Psychiatria Hungarica, 16(4), 379–397.
Sadaghiani, S., Scheeringa, R., Lehongre, K., Morillon, B., Giraud, A.-L., & Kleinschmidt, A. (2010). Intrinsic connectivity networks, alpha oscillations, and tonic alertness: A simultaneous electroencephalography/functional magnetic resonance imaging study. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience, 30(30), 10243–10250.
Schredl, M. (2009a). Nightmare frequency in patients with primary insomnia. International Journal of Dream Research, 2(2), 85–88.
Schredl, M. (2009b). Dreams in patients with sleep disorders. Sleep Medicine Reviews, 13(3), 215–221. Schredl, M. (2010). Nightmare frequency and nightmar
e topics in a representative German sample. European Archives of Psychiatry and Clinical Neuroscience, 260(8), 565–570.
Schredl, M., Schafer, G., Weber, B., & Heuser, I. (1998). Dreaming and insomnia: Dream recall and dream content of patients with insomnia. Journal of Sleep Research, 7(3), 191–198.
Schürmann, M., & Bas¸ar, E. (2001). Functional aspects of alpha oscillations in the EEG. International Journal of Psychophysiology: Official Journal of the International Organization of Psychophysiology, 39(2–3), 151–158.
Simor, P., Bódizs, R., Horváth, K., & Ferri, R. (2013). Disturbed dreaming and the instability of sleep: Altered NREM sleep microstructure in subjects with frequent nightmares as revealed by the cyclic alternating pattern. Sleep, 36(3), 413–419.
Simor, P., Horváth, K., Gombos, F., Takács, K. P., & Bódizs, R. (2012). Disturbed dreaming and sleep quality: Altered sleep architecture in subjects with frequent nightmares. European Archives of Psychiatry and Clinical Neuroscience, 262(8), 687–696.
Simor, P., Köteles, F., Bódizs, R., & Bárdos, G. (2009). A szubjektív alvásminőség kérdőíves vizsgálata: A Gröningen Alvásminőség Skála hazai validálása. Mentálhigiéné és Pszichoszomatika, 10(3), 249–261.
Simor, P., Kovács, I., Vargha, A., Csóka, S., Mangel, B., & Bódizs, R. (2009). Nightmares, dream anxiety and psychopathology: The validation of the Hungarian version of the Van Anxiety Scale. Psychiatria Hungarica: A Magyar Pszichiátriai Társaság Tudományos Folyóirata, 24(6), 428–438.
Sipos, K., Sipos, M., & Spielberger, C. (1994). A state-trait anxiety inventory (STAI) magyar változata. Pszichodiagnosztikai Vademecum, 2.
Spielberger, C., Gorsuch, R., & Lushene, R. (1970). Manual for the STAI. Consulting Psychologists Tests.
Spiegelhalder, K., Regen, W., Feige, B., Holz, J., Piosczyk, H., Baglioni, C., et al. (2012). Increased EEG sigma and beta power during NREM sleep in primary insomnia. Biological Psychology, 91(3), 329–333.
Spoormaker, V. I., Schredl, M., & van den Bout, J. (2006). Nightmares: From anxiety symptom to sleep disorder. Sleep Medicine Reviews, 10(1), 19–31.
Steriade, M., & Llinás, R. R. (1988). The functional states of the thalamus and the associated neuronal interplay. Physiological Reviews, 68(3), 649–742.
Steriade, M., McCormick, D. A., & Sejnowski, T. J. (1993). Thalamocortical oscillations in the sleeping and aroused brain. Science (New York, N.Y.), 262(5134), 679–685.
Taylor, F., & Raskind, M. A. (2002). The alpha1-adrenergic antagonist prazosin improves sleep and nightmares in civilian trauma posttraumatic stress disorder. Journal of Clinical Psychopharmacology, 22(1), 82–85.
Terzano, M. G., Mancia, D., Salati, M. R., Costani, G., Decembrino, A., & Parrino, L. (1985). The cyclic alternating pattern as a physiologic component of normal NREM sleep. Sleep, 8(2), 137–145.
Tribl, G. G., Wetter, T. C., & Schredl, M. (2012). Dreaming under antidepressants: A systematic review on evidence in depressive patients and healthy volunteers. Sleep Medicine Reviews, 17(2), 133–142.
Tyson, P. D., Ogilvie, R. D., & Hunt, H. T. (1984). Lucid, prelucid, and nonlucid dreams related to the amount of EEG alpha activity during REM sleep. Psychophysiology, 21(4), 442–451.