|
DOI: 10.1556/MPSzle.63.2008.1.4 Semmelweis Egyetem, Magatartástudományi Intézet Beérkezett: 2007. 12. 10. – Elfogadva: 2008. 01. 15. *A szerzőt a Magyar Tudományos Akadémia Bolyai János Kutatói Ösztöndíjjal támogatja. AbsztraktA neurális rendszerek szinkronizált oszcillációkba szerveződése az alvás elektrofiziológiailag jól megragadható vonatkozásainak egyike. Az oszcillációk általános, mindenkire jellemző jegyei mellett egyre ismertebbé válnak a személyre specifikus vonások is, amelyek az alvás egyéni topografikus és spektrális electroencefalográfiás (EEG) ujjlenyomatai formájában számszerűsíthetők. Az egyéni alvás-EEG ujjlenyomatokra a figyelemre méltó időbeli stabilitás, az idegrendszer plasztikus folyamataival való kétirányú kapcsolat és a neurális konnektivitás általi nagyfokú meghatározottság jellemző. Ez utóbbi következtében az alvás-EEG ujjlenyomatok alkalmasak lehetnek az egyének közötti pszichológiai különbségek neurobiológiai vonatkozásainak kutatására. E megközelítés helytállóságát egyre több bizonyíték támogatja. Az alvásra jellemző lassú hullámú aktivitás összefüggésben áll a memória és a végrehajtó funkciók hatékonyságával, míg az alvási orsók és a kognitív teljesítmény között bonyolult sokrétű kapcsolatrendszer feltételezhető, amiben az általános mentális rátermettség vagy az intelligencia neurobiológiája is új megvilágításba kerülhet. Az affektív szféra és az alvás-EEG korrelációja tekintetében rendelkezésre álló szórványos adatok szintén az alvás-EEG differenciálpszichológiai jelentőségét erősítő megfigyelések. Az alvás-EEG oszcillációk az egyéni funkcionális neuroanatómiától való függésük következtében tükrözik a neurális kapcsolatrendszer jelenlegi állapotát, de a plasztikus folyamatok előtérbe helyezése révén egyben lehetőséget is teremtenek a rendszer hatékony és adaptív újjászervezésére. Kulcsszavak: alvás-EEG ujjlenyomatok, konnektivitás, intelligencia, kognitív képességek, memória, alvási orsók, delta hullámok, lassú oszcilláció SLEEP PHYSIOLOGY AS AN APPROACH OF DIFFERENTIAL PSYCHOLOGY: SIGNIFICANCE OF INDIVIDUAL EEG PATTERNSOne of the well described aspects of the electrophysiology of sleep is related to the organization of neural systems in synchronized oscillations. Beside the general aspects of these oscillations there is increasing evidence for the existence of some individual-specific traits, which can be quantified by the individual topographic and spectral fingerprints of sleep-EEG. Individual fingerprints of human sleep-EEG are characterized by remarkable stability, bidirectional relationship with neural plasticity and definiteness by neural connectivity. Given the latter characteristic, these fingerprints could play an important role in research work aiming to reveal the neurobiological bases of psychological differences. Accumulating evidence supports the pertinence of this approach. Correlations of sleep-dependent slow wave activity with memory and executive functions were found, while sleep spindling was shown to have a complex relationship with cognitive performance. This relationship may shed light on the neurobiological bases of general mental ability or intelligence. Some preliminary data supporting the correlation between sleep-EEG and the affective traits of human subjects strengthen the differential psychological relevance of sleep-EEG activity. Given the fact that individual functional neuroanatomy is a major factor that shapes sleep-EEG oscillations, the latter reflect the actual state of synaptic infrastructure. However, by facilitating plastic processes neural oscillations also open the door to the efficient and adaptive reorganization of neural connectivity. Key words: sleep-EEG fingerprints, connectivity, intelligence, cognitive abilities, memory, sleep spindles, delta waves, slow oscillation ELMÉLETI MEGFONTOLÁSOKAz egyének közötti különbségek kutatása hosszú múltra tekint vissza, a tudományos diszciplínák igen széles skáláját ölelve fel. Ezen a skálán a molekuláris genetikától a szociológiáig és az antropológiáig az elemzések eltérő szintjeit és a megközelítésmódok széles spektrumát találjuk. Mindazonáltal a kultúra, a szociális tényezők és a genetika személyiséget létrehozó és alakító hatásainak egyfajta végső közös útjának az idegrendszer egyéni felépítése tekinthető (DAVIDSON, 2001). Az egyéni különbségek vizsgálatának és becslésének számottevő tudományos és társadalmi jelentőségére számos megfigyelés utal (LUBINSKI, 2000). Ennek ellenére — néhány jelentős kezdeményezéstől (GRAY, THOMPSON, 2004) eltekintve — még mindig igen szegényesnek, szervezetlennek és szórványosnak mondható azon tudományos kutatási programok köre, amelyek kifejezetten a viselkedésbeli különbségek idegrendszeri korrelátumait kívánják feltárni. Az idegtudományok jelenlegi fő árama a viselkedéses és lélektani jelenségek olyan közös neurális mechanizmusainak föltárásával foglalkozik, amelyeknek eredményeképpen általános, mindenkire érvényes modellek szület(het)nek. A részmechanizmusok egyre mélyebb feltárása azonban háttérbe szorította a stabil személyi sajátosságok mögötti biológia megismerését, azaz a hangsúlyok rendszerint az időbeli lefolyáson és a hatásokon vannak a stabil mutatók és a vonások helyett, ami a biológiai rendszerek egyenlő alakíthatóságának mögöttes metaelméletét implikálja. Az egyéni különbségek problémakörének mellőzöttségéhez burkoltan az a jelenleg érvényes kutatási hagyomány is hozzájárul, miszerint a vizsgálatokat mesterségesen és erőteljesen homogenizált, korántsem reprezentatív (leggyakrabban egyetemi hallgatókból álló) populációkon kell végezni. Az egyéni különbségek kutatása – a genetikától eltekintve – a közhelyek szintjén van, és kívül maradt napjaink idegtudományának, illetve pszichológiájának fő áramán. Saját kutatási programunk és egyben a jelenlegi összefoglaló célja az egyéni különbségek idegtudományának kiegészítése egy új témával: az alvás-electroencefalogram (alvás-EEG) egyéni mintázatainak differenciálpszichológiai jelentőségével. Hangsúlyozni kívánjuk, hogy a viselkedéses fenotípusok funkcionális és neuroanatómiai alapjainak feltárásában és megértésében kiemelkedő szerep jut az alváskutatásnak. Ugyanakkor – a dimenzionális megközelítést követve – feltételezhető, hogy a különböző pszichopatológiai és neuropszichiátriai jelenségeket, mint a kognitív és affektív sajátosságok extrém megnyilvánulásait eddig ismeretlen biológiai markerekkel társítjuk, illetve új mechanizmusszintű és konceptuális értelmezési keretekkel magyarázzuk egy ilyen megközelítésmód sikeres alkalmazása esetén. AZ ALVÁS-EEG EGYÉNI UJJLENYOMATAIAz állapotok és a vonások lényegi különbsége az időbeli változás, illetve stabilitás tulajdonságaiban keresendőek. Amennyiben az egyéni különbségek kutatását az alvással kapcsolatos fiziológiai paraméterekre alapozzuk, alapvető feltétel, hogy a megfelelő paraméter időben stabil legyen. Ez a sarkalatos kérdés elvileg minden a személyiség, illetve a kognitív képességek terén végzett pszichofiziológiai kutatással kapcsolatban fölmerül. Az alábbiakban az alvás-EEG némely karakterisztikus vonásának időbeli állandóságával, valamint változatosságával kapcsolatos kutatás kritikai áttekintését adjuk meg. Lassú hullámú EEG-tevékenységAz alvás-EEG egyik lényegi vonása a lassú hullámú tevékenység (<4 Hz) ébrenléthez viszonyított dominanciája (BÓDIZS, 2000). A szakirodalom jelentős része foglalkozik az alvás alatti lassú hullámú EEG-tevékenység homeosztatikus szabályozásával, ami közvetve az alvás homeosztatikus szabályozását tükrözi. Ez azt jelenti, hogy a jellemzően a nem-REM (NREM) alváshoz kötötten megjelenő lassú hullámú EEG-tevékenység mértéke, az alvást megelőzően ébren töltött idővel, illetve regionálisan, a központi idegrendszeri (többnyire, de nem kizárólag kérgi) struktúrák használatával arányos (BORBÉLY, 2001). Ez a változékonyság elvileg kizárná a vonásszerűség elvének alkalmazását a lassú hullámú EEG-tevékenységgel kapcsolatosan, azonban egyes újabb eredmények más következtetéseket is megengednek, sőt úgy tűnik, valószínűbbé is tesznek. A lassú hullámú EEG-tevékenység teljesítménysűrűség-spektrumának ismétlési megbízhatóságával kapcsolatos korrelációs vizsgálatok közül az egyik legalaposabb 0,84-es erősségű összefüggésről számol be, ami azt jelenti, hogy az éjszakai alvás ezen EEG-mutatója 70%-os pontossággal előre jelezhető egyszerűen csak egy korábbi éjszakai vizsgálat eredményének ismeretében (TAN, CAMPBELL és munkatársai, 2000). Ez elvileg azt is jelentheti, hogy a személyek megbízhatóan különböznek napközbeni aktivitásuk és ébrenléti idejük tekintetében, ami az éjszakai lassú hullámú EEG-tevékenységben is tükröződik. Egy újabb vizsgálatban azonban az alvásmegvonás hatását is figyelembe véve kimutatták, hogy az alvás alatti lassú hullámú EEG-tevékenység egyének közötti változatosságának foka majdnem tízszerese annak a változásnak, amit egy éjszakai alvás kimaradása révén az ébrenlét idejének meghosszabbodása okoz (TUCKER, DINGES, VAN DONGEN, 2007). Az egyének közötti különbségek tehát az ébren töltött időtől függetlenül is igen jelentősek, sőt a korábbi kutatások elsőrendű fókuszával ellentétben jelentősebbeknek mondhatóak, mint az amúgy kimutatható és funkcionálisan jelentős homeosztatikus alvásszabályozás. Ezek az eredmények összhangban vannak egyes korábbi ikervizsgálatokkal. Utóbbiak szerint ugyanis számottevő genetikai meghatározottság mutatható ki az alvás lassú hullámokkal jellemezhető szakaszainak (3. és 4. stádiumok) időtartama tekintetében, ami közvetve a fenti megfigyeléseket erősítő adat (LINKOWSKI, KERKHOFS és munkatársai, 1989, 1991). Újabb ikervizsgálatok ugyanakkor arra hívják fel a figyelmet, hogy a lassú hullámú EEG-tevékenység időbeli lefutásának mintázatában is erőteljes genetikai hatások érvényesülhetnek (KIMURA, WINKELMAN, 2007). A lassú hullámú EEG-tevékenység térbeli (skalp fölötti) eloszlása tekintetében hasonló eredmények születtek. A skalp fölötti eloszlás mintázatát nagyfokú egyéni változatosság és kimagasló személyeken belüli stabilitás jellemzi. Az alvásmegvonást követő alvásban mért lassú hullámú EEG-tevékenység fokozott ugyan az alvásmegvonás előttihez képest, mégis a normalizált spektrális értékek térképe változatlan és egyénre jellemzően stabil marad (FINELLI, ACHERMANN, BORBÉLY, 2001). A hasonló eredmények robusztussága vezette Luca Finellit és munkatársait (2001) arra, hogy az alvás EEG-topográfiájának egyéni ujjlenyomatairól beszéljen, ami szerintük az idegrendszer egyéni funkcionális neuroanatómiáját tükrözi. Alvási orsózás és alvás alatti alfa aktivitásAz alvási orsók hozzávetőlegesen 12–15 Hz-es (más meghatározások szerint 9–16 Hz-es) szinuszoid hullámcsomagok, amelyeknek a kezdeti növekvő, majd fokozatosan csökkenő amplitúdó orsóalakot kölcsönözhet (1. ábra). Leggyakrabban a 2-es stádiumú alvásban figyelhetőek meg, de a NREM alvás minden fázisában jelen lehetnek (DE GENNARO, FERRARA, 2003).
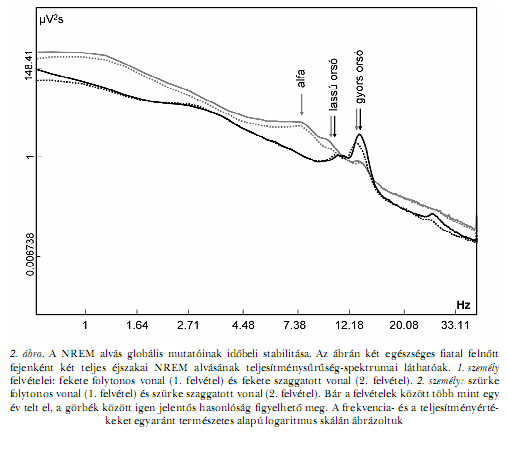
Az alvási orsókra jellemző frekvenciasávban az EEG-tevékenység teljesítménysűrűség-spektrumának ismétlési megbízhatósága még a lassú hullámú tevékenységénél is kimagaslóbb, TAN, CAMPBELL és munkatársai (2000) vizsgálati eredményei szerint 0,98. Az orsóknak megfelelő frekvenciatartományú EEG-teljesítménysűrűségspektrumok skalp fölötti megoszlásának mintázatára ugyanaz az ujjlenyomatszerűség jellemző, mint a lassú hullámú tevékenységére (FINELLI, ACHERMANN, BORBÉLY, 2001). Az orsók spektrális és topográfiai jellemzői azonban összetettebbek a lassú hullámú tevékenységénél. Az emberek nagyobb részében két elkülönült spektrális összetevő (csúcs) figyelhető meg az orsófrekvencián belül (2. ábra). Ezek topografikusan is különböznek: a lassúbb csúcsnak (~11 Hz) megfelelő EEG-tevékenység frontális maximumú, míg a gyorsabb csúcsnak megfelelő tevékenység parietális maximumú. A lassú és a gyors orsózás agyi hemodinamikai sajátosságainak vannak közös és eltérő elemei. Mindkét orsótípust a thalamus, az elülső cinguláris kéreg, az insularis kéreg és a superior temporalis gyrus fokozottabb agyi vérátáramlása jellemzi. Ugyanakkor a superior frontalis gyrus elsősorban a lassú orsók ideje alatt, míg a szenzomotoros területek, a temporális lebeny mediális felszíne és a hippocampus a gyors orsók ideje alatt aktívabbak (SCHABUS, DANG-VU és munkatársai, 2007). A kétféle orsófrekvenciának megfelelő spektrális csúcs egymáshoz viszonyított dominanciája, pontos frekvenciája és topográfiája egyénre jellemző stabil tulajdonság (WERTH, ACHERMANN és munkatársai, 1997; DE GENNARO, FERRARA és munkatársai, 2005). Az alfa hullámoknak és az alvási orsózásnak megfelelő frekvenciaövezet (8–15 Hz) vonás-, illetve állapotjellemzőinek részletes vizsgálata nyomán kiderült, hogy bár mindkét orsótípusban kimutathatóak állapottól függő hatások (alvásmegvonás vagy az alvás folyamatának kísérletes célú megzavarása), ennek ellenére az EEG teljesítményspektrumának z-transzformált értékei kiemelkedően stabil személyi jellemzők, melyeknek ismétlési megbízhatósága minden esetben 0,90 fölötti (DE GENNARO, FERRARA és munkatársai, 2005). Ugyanakkor az orsófrekvenciájú EEG-aktivitás időbeli lefutásának mintázata egy ikervizsgálat előzetes eredményei szerint számottevő genetikai hatások tükrözője is egyben (KIMURA, WINKELMAN, 2007). Fontos megjegyezni, hogy az orsófrekvenciájú EEG-aktivitás spektruma nem tükrözi tökéletesen a jellegzetes fázisos orsótevékenységet, ami időben elkülönülő EEG-jelenségek ismétlődéséből áll. A teljesítményspektrum ugyanis olyan EEG-komponensekre is érzékeny, amelyek nem csoportosulnak orsókká, de orsófrekvenciájúak (DE GENNARO, FERRARA, 2003). Ezért az orsók detekciója és az EEG- tevékenység teljesítményspektrumának orsófrekvenciájú összetevői nem teljesen azonosak. Mindazonáltal a vizuálisan és az automatikusan detektált orsók száma és időbeli megoszlásának mintázata szintén stabil személyi jellemzőnek bizonyult (SILVERSTEIN, LEVY, 1976; AZUMI, SHIRAKAWA, 1982; GAILLARD, BLOIS, 1981). Az alvás-EEG globális mutatóinak stabilitása és személyfüggőségeRészben az eddig említett eredmények által indíttatva több próbálkozás is történt a globális, teljes frekvenciatartományra kiterjedő alvás alatti EEG-tevékenység vonásjellegének tanulmányozására. Az egyik, erre a kérdésre összpontosító vizsgálat eredményei szerint a húsznapos időközönként fölvett, személyenként négy alvás-EEG logaritmizált teljesítménysűrűség-spektrumai majdnem tökéletes egyezést mutatnak a teljes frekvenciatartományon, NREM és REM alvásban, illetve fiataloknál és időseknél egyaránt. Ugyanakkor megfigyelhető egy jelentős életkori hatás is (a fiatal és az idős csoport közötti szignifikáns különbségekkel), tehát az időbeli változatlanság bizonyos keretek között kezelendő (TAN, CAMPBELL, FEINBERG, 2001). Saját adatbázisunkban rendelkezésre álló több mint egy éves utánkövetéses adatok feldolgozása nyomán is rendkívül figyelemreméltó időbeli stabilitás látható (2. ábra). Egy újabb vizsgálatban 8 személy, fejenként négy teljes éjszakai alvás-EEG spektrumából matematikai eljárással vonásvektorokat képeztek, majd ezeket klaszteranalízisnek vetették alá. A klaszteranalízis éppen 8 klasztert mutatott ki, amelyek a 8 vizsgált személy klaszterenként 4 felvételét tartalmazták. Vagyis minden személy nagyobb mértékben hasonlít saját korábbi felvételeihez, mint másokéhoz (BUCKELMÜLLER, LANDOLT és munkatársai, 2006). Egy ikervizsgálat előzetes eredményei arra utalnak, hogy az alvás alatt rögzített EEG-felvételek teljesítményspektrumainak alakjában jelentős genetikai hatások is érvényesülhetnek (KIMURA, WINKELMAN, 2007).
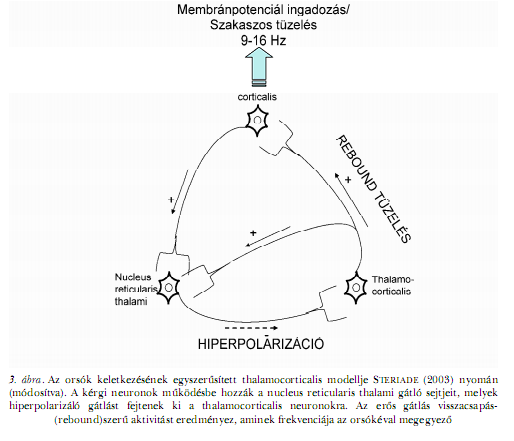
ALVÁS-EEG, KONNEKTIVITÁS, IDEGRENDSZERI PLASZTICITÁSKonnektivitás és plaszticitás a lassú hullámok viszonylatábanEgyre több bizonyíték támogatja a NREM alvás alatti lassú hullámú EEG- tevékenységnek a kérgi szinaptikus háló szerkezeti és működési sajátosságaival való kapcsolatát. Az állatkísérletes adatok egyike a korai vizuális depriváció paradigmáját alkalmazta a látókéreg szerkezetének befolyásolása céljából. A szerzők kimutatták, hogy a sötétben nevelt állatok látókérge fölött kevésbé kifejezett az alvás alatti lassú hullámú EEG-aktivitás (MIYAMOTO, KATAGIRI, HENSCH, 2003). A szerzők kimutatták, hogy a folyamat az NMDA-receptorok aktiválásával áll kapcsolatban, és az idegrendszeri plaszticitás egyik megnyilvánulási formája. A szinaptikus plaszticitás és az alvásfüggő lassú hullámú tevékenység között kétirányú kapcsolat figyelhető meg (MIYAMOTO, HENSCH, 2006), ami azért is újszerű meglátás, mert rámutathat az embereknél megfigyelt stabil egyéni különbségek természetére, a használattól függő szinaptikus plaszticitás mechanizmusa révén. Korábbi kísérleti adatokban is találhatunk bizonyítékokat arra vonatkozóan, hogy az alvással összefüggő lassú hullámú EEG-tevékenység tükrözi a szinaptikus jelátvitel hatékonyságát. AMZICA és STERIADE (1995) kísérletében macskák két távoli kérgi területe között helyi érzéstelenítőkkel felfüggesztették a szinaptikus jelátvitelt. Ennek hatására a két terület 1 Hz alatti elektromos oszcillációja, a lassú oszcilláció, aszinkron módon jelent meg, holott ennek a tevékenységnek a szinkronizáció alapvető tulajdonsága. ESSER, HILL és TONONI (2007) modellkísérletben mutattak rá arra, hogy a kérgi szinaptikus jelátvitel gyengülése csökkenti a lassú hullámok megjelenésének gyakoriságát az EEG-ben. Ezt olyan változások kísérték, mint a hullámok meredekségének csökkenése és a jól kivehető csúcsokkal jellemezhető lassú hullámok helyett felbukkanó többszörös csúcsokat mutató lassú hullámok. A lassú hullámú EEG-tevékenységen belül elkülöníthető egy lassú oszcillációnak nevezett hullámtevékenység és az úgynevezett delta aktivitás. A lassú oszcilláció frekvenciája 1 Hz alatti, és az egyedi kérgi neuronokban, illetve neuronhálózatokban keletkezik. Terjedése és szinkronizációja szinte teljes mértékben az intrakortikális konnektivitás útján (tehát hatékony, működő szinapszishálózatok révén) valósul meg (STERIADE, 2003). Ezek a neurofiziológiai eredmények tehát arra utalnak, hogy maga a lassú oszcilláció egyben a konnektivitás mutatója is, a konnektivitás pedig a használattól függő és a genetikailag meghatározott plaszticitás eredője (BÓDIZS, 2006). Ugyanakkor, az alvási orsók triggerelése révén közvetve maga a lassú oszcilláció is elősegíti a plaszticitást (STERIADE, 2006). Egyes megfigyelések szerint a delta hullámok, melyeknek frekvenciája hozzávetőlegesen 1 és 4 Hz közötti – a lassú oszcillációval szemben – legalábbis részben, a thalamus és a kéreg közötti kapcsolatrendszerben keletkeznek (STERIADE, 2003), alvásciklusok közötti dinamikájuk, illetve homeosztatikus alvásszabályozásban játszott szerepük pedig eltér a lassú oszcillációétól (ACHERMANN, BORBÉLY, 1997; CAMPBELL, HIGGINS és munkatársai, 2006). Míg egyes kognitív vizsgálatok eredményei szerint jogos a lassú oszcilláció és a delta hullámok megkülönböztetése (BÓDIZS, BÉKÉSY és munkatársai, 2002; ANDERSON, HORNE, 2003), más vizsgálatok szerint a két szóban forgó frekvenciatartomány funkcionálisan egységesnek tekinthető (MIYAMOTO, KATAGIRI, HENSCH, 2003; ESSER, HILL, TONONI, 2007; MASSIMINI, FERRARELLI és munkatársai, 2007). Konnektivitás és plaszticitás az alvási orsók viszonylatábanA celluláris neurofiziológiai vizsgálatok eredményeinek fényében feltételezhető, hogy az alvás-EEG orsózó hullámformái a thalamocorticalis neuronok ritmikus hiperpolarizáció-rebound szekvenciáinak a kérgi neuronok membránpotenciáljaiban való tükröződései, amelyekben a nucleus reticularis thalami neuronjai képezik a gátló komponenst (STERIADE, 2003; 2006). Az orsók keletkezési mechanizmusának ezen modellje szerint a thalamocorticalis hurkok az orsózás funkcionális elemei (3. ábra).
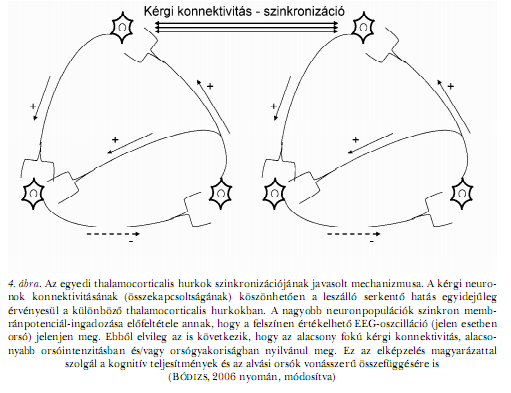
Mindez azonban feltételez egy szinkronizáló mechanizmust is, ami az egyedi thalamocorticalis hurkokban egyidejűvé teszi az oszcillációt. Tekintve, hogy a thalamuson belüli kapcsolatok szegényesek, ezt a szinkronizáló hatást leginkább a kéreg biztosíthatja, az intrakortikális konnektivitás és a lassú oszcilláció révén. Feltételezhető, hogy a lassú oszcilláció neuronálisan aktív szakaszában a leszálló corticothalamicus rostok szinkron módon hozzák működésbe a thalamus orsózó üzemmódját (STERIADE, 2006). Ennek a mechanizmusnak köszönhető, hogy az orsózás spontán módon a lassú oszcilláció meghatározott fázisaiban (a felszínen regisztrált pozitív fázisban) csúcsosodik ki. Ezt hívják az alvási orsók lassú oszcilláció általi csoportosításának (MÖLLE, MARSHALL és munkatársai, 2002). Vagyis az igen nagyszámú kérgi neuron membránpotenciál-ingadozásának összegződéséből kirajzolódó felszíni EEG-ben nem is láthatnánk orsókat, ha a kérgi szinkronizáló hatás nem érvényesülne, illetve a lassú oszcilláció nem csoportosítaná az orsózást. Ebből elvileg az is következik, hogy a kérgi kapcsolatrendszer/konnektivitás hatékonysága az orsózás élénkségében is kifejeződik (BÓDIZS, 2006; 4. ábra).
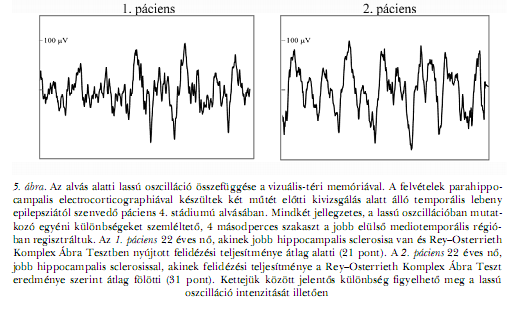
Mindemellett az orsózásnak közvetlen hatása is lehet az idegrendszeri plaszticitásra. A fokozódó válasz (augmenting response) jelensége régről ismert az idegélettanban. Lényege, hogy a thalamus felszálló rostjainak 8–15 Hz-en történő ritmikus ingerlése közben a kérgi válasz fokozatosan nő, anélkül hogy az ingerlés erősségét változtatnánk (MAGOUN, 1963). Ez a frekvencia az alfa hullámok és az alvási orsók tartományával megegyező. Mára ismertté vált, hogy a spontán megjelenő alvási orsók ideje alatti egyszeri (nem ritmusos) kérgi ingerlés sokkal tartósabb nyomot hagy az ingerelt neuronpopulációk válaszkészségében, mint a nem orsózó szakaszban adott ingerlés (TIMOFEEV, GRENIER és munkatársai, 2002). Számos hasonló megfigyeléssel párosítva ezek az eredmények arra utalnak, hogy az alvási orsózásnak megfelelő agyi üzemmód (8–15 Hz-es ritmikus tüzelés) fokozottan plasztikus állapot, amelyben fokozott az esélye a neuronális hálózatok módosulásának és átszerveződésének. Vagyis az alvási orsók vélhetően egyszerre tükrözik és formálják a kérgi, illetve corticothalamicus kapcsolatrendszert. ALVÁS-EEG ÉS KOGNITÍV TELJESÍTMÉNYLassú hullámú EEG-aktivitás alvásban és kognitív teljesítmény ébrenlétben: figyelemre méltó összefüggésekAz alvás NREM fázisában regisztrált lassú hullámú EEG-tevékenység bizonyos mutatói és az ébrenlét állapotában becsült kognitív teljesítmények különböző összetevői közötti kapcsolat gondolata nem új a szakirodalomban. A kutatók nagyobbik része elsősorban az alvásnak a kognitív tevékenységekre gyakorolt kedvező hatása keretében vizsgálja, illetve értelmezi az eredményeit. Anélkül, hogy tagadnánk az alvásnak a kognitív teljesítményekre gyakorolt hatását, ehelyütt elsősorban azokat a vonatkozásokat igyekszünk kidomborítani, amelyek a két jelenség közötti vonásszerű kapcsolatra engednek következtetni. A rendelkezésre álló adatok egy része azt a megállapítást látszik támogatni, miszerint az alvás NREM fázisában regisztrált lassú oszcilláció (< 1 Hz) kifejezett volta a kognitív teljesítmények kedvező mutatójának tekinthető. Az egészséges idős személyek alvásának első NREM fázisában a frontális területek fölött mért lassú oszcilláció relatív spektrális teljesítménye pozitívan korrelált a végrehajtó funkciók becslésére irányuló neuropszichológiai tesztekben nyújtott teljesítményekkel (ANDERSON, HORNE, 2003). Utóbbira példa a verbális fluencia becslésének egyik változata (a vizsgálati személy azon képessége, hogy egy-egy igéhez értelmesen rendeljen hozzá minél több főnevet) vagy a London-torony megoldásának képessége. ANDERSON és HORNE (2003) az alvásvizsgálat után egy héttel tesztelték a neuropszichológiai funkciókat, mégis fölvetik az alvás proaktív hatásának lehetőségét is (az alvás ideje alatti lassú hullámú EEG-tevékenység életkorfüggő csökkenésével egyetemben). E tekintetben fontos ismét megemlíteni, a lassú hullámú EEG-tevékenység tekintetében mért egyének közötti különbség meglehetősen stabil és jelentős voltát (TUCKER, DINGES, VAN DONGEN, 2007). Ennek fényében pedig az oksági láncolatról nem sokat mondhatunk egy korrelatív vizsgálat eredményeinek értelmezésekor. Vagyis a jól teljesítő személyek frontális lassú oszcillációjának erőteljessége éppen úgy lehet a habituális napközbeni igénybevétel következménye, mint annak az előidézője, vagy akár egy harmadik közös – a teljesítményt és az EEG-t egyaránt befolyásoló – tényező eredője. Saját korábbi vizsgálataink egyikében epilepsziás populáción mutattuk ki az intrakraniális elektródákkal regisztrált temporo-mediális és a hagyományos felszíni elektródákkal mért temporo-laterális lassú oszcilláció relatív teljesítménye és a Rey–Osterrieth Komplex Ábra felidézése (emlékezetből történő újrarajzolása) közötti szoros összefüggést, úgy, hogy az alvás és a neuropszichológiai tesztelés között hetek, esetenként hónapok teltek el (BÓDIZS, BÉKÉSY és munkatársai, 2002). Bár az eredményeket az epilepszia és minden bizonnyal az antiepileptikus gyógyszeres kezelés is befolyásolhatta, mégis figyelemre méltó, hogy a klinikailag igen hasonlónak látszó esetek közötti különbség szemmel is megfigyelhető alvás- EEG különbségekkel társult (5. ábra). Magyarul az alvás-EEG jobb előrejelzője volt a teljesítménynek, mint a klinikai státusz.
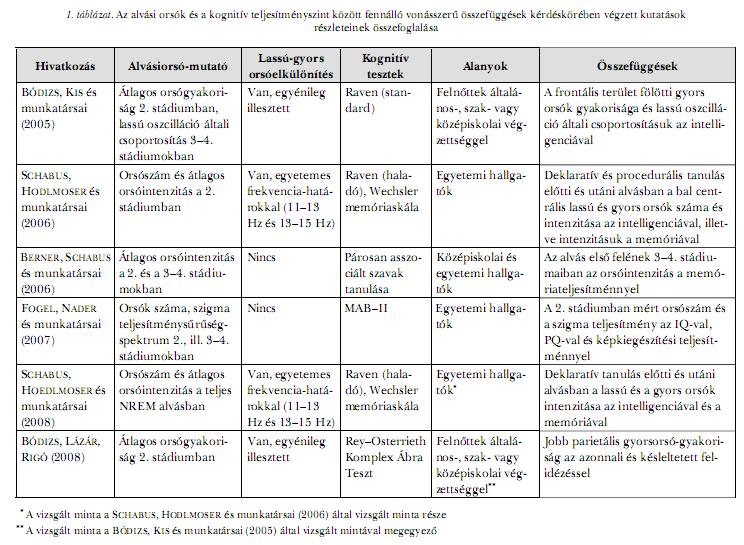
Egy újabb vizsgálat eredményei szerint a 3–4. stádiumú alvás-EEG teljes lassú hullámú tartományának (0,5–4 Hz) logaritmizált teljesítménysűrűség-spektruma pozitívan korrelál nemcsak a Rey–Osterrieth Komplex Ábra Tesztben, de a Trailmaking Tesztben nyújtott teljesítménnyel is. Erőteljes összefüggéseket elsősorban a frontális régióban figyeltek meg (GÖDER, ALDENHOFF és munkatársai, 2006). Stabil amisulprid-kezelés alatt álló szkizofrén populációban szignifikánsan csökkent a lassú hullámú alvás (3–4. stádiumok) ideje, ami a Rey–Osterrieth Komplex Ábra Tesztben nyújtott teljesítményromlás előrejelzőjének bizonyult (GÖDER, BOIGS és munkatársai, 2004). Csökkent a 3–4. stádiumok lassú hullámú EEG- tevékenységének teljesítménysűrűség-spektruma is, ez viszont – a kontrollmintával ellentétben – nem korrelált a Rey–Osterrieth Komplex Ábra Tesztben nyújtott teljesítményekkel, csak a Trail Making pontszámokkal (GÖDER, ALDENHOFF és munkatársai, 2006). Összességében elmondható, hogy az eddigi megfigyelések értelmében az alvás alatt mért lassú hullámú EEG-tevékenység egyéni eltérései elsősorban a kivitelező funkciókkal vagy általában a homloklebenyi működésekre érzékeny feladatokkal és a vizuális-téri emlékezeti teljesítmény egyéni különbségeivel állhatnak kapcsolatban. Alvási orsók és kognitív teljesítményAz alvási orsók és a kognitív teljesítmény közötti kapcsolat jelenleg intenzív kutatások tárgya. A vizsgálati eredmények döntő többségének preferált értelmezési kerete az alvási orsóknak az emlékezeti rögzülésben játszott szerepével kapcsolatos. Ezzel kapcsolatban igen sokrétű empirikus bizonyíték látott napvilágot (CLEMENS, FABÓ, HALÁSZ, 2005; 2006; SCHABUS, HODLMOSER és munkatársai, 2008; GAIS, MÖLLE és munkatársai, 2002). Ugyanakkor jelentős megfigyelések utalnak arra is, hogy az orsózásban mutatkozó egyének közötti különbségek kapcsolatban állnak a kognitív képességekkel (BÓDIZS, KIS és munkatársai, 2005; FOGEL, NADER és munkatársai, 2007; BERNER, SCHABUS és munkatársai, 2006; BÓDIZS, LÁZÁR, RIGÓ, 2008). A két hatás együttes értelmezésére lehetőséget adó vizsgálati eredmények újabban szintén felbukkantak e témakörben (SCHABUS, HODLMOSER és munkatársai, 2006; 2008). Az alvási orsózás egyéni szintjének különböző mutatói és az intelligencia közötti kapcsolat az egyik e témában közölt jelentős megfigyelés. Az alvás 2. stádiumában a frontális területek fölött mért percenkénti gyorsorsó-gyakoriság és a Raven Progresszív Mátrix Tesztben nyújtott teljesítmény közötti – életkorra korrigált – pozitív korreláció az egyik eredmény. A 3–4. alvásstádiumokban mért lassú oszcilláció általi gyorsorsó-csoportosítás és a Raven Tesztben nyújtott teljesítmény összefüggése szintén prefrontális lokalizációjú (BÓDIZS, KIS és munkatársai, 2005). Hasonló eredményt közöltek az orsózás átlagos erőssége tekintetében: az orsók átlagos amplitúdójának és időtartamának szorzata pozitív kapcsolatban állt a Raven Progresszív Mátrix Teszt haladó változatában nyújtott teljesítménnyel egyetemi hallgatók körében. Az összefüggés a lassú és a gyors orsókkal kapcsolatban is szignifikáns volt, és igen kevéssé függött attól, hogy alvás előtt a vizsgált személyek procedurális vagy deklaratív anyag elsajátításában vettek-e részt vagy nem kaptak tanulásra utaló instrukciót, illetve feladatot (SCHABUS, HODLMOSER és munkatársai, 2006; 2008). FOGEL, NADER és munkatársai (2007) a lassú és a gyors orsók differenciált elemzése nélkül vizsgálták az orsók gyakorisága, valamint az orsófrekvenciájú EEG-aktivitás teljesítményspektruma és a MAB–II (Multiple Aptitude Battery) intelligenciateszt eredményei közötti összefüggést. Mindkét orsó-mutató 2. stádiumú alvásra számított értéke pozitívan korrelált az intelligenciahányadossal és a performációs (cselekvéses) hányadossal (PQ), de nem függött össze a verbális teljesítménnyel (VQ). A legerősebb kapcsolatot az alvási orsózás és a képkiegészítés próba között találták. Ugyanakkor fölmerült egy arra utaló eredmény is, amely szerint az összefüggés nem minden intelligenciaövezetben érvényes, és esetleg csak a 115 és 126 közötti PQ értékű személyek körében mutatható ki. Az alvási orsózásban megfigyelhető egyének közötti különbségek azonban nemcsak az intelligenciatesztekben nyújtott kognitív teljesítményekkel, hanem más, specifikusabb képességekkel is kapcsolatban állhatnak, ami némileg ellentmondásossá, de mindenképpen összetettebbé teszi a képet. Bár a párosan asszociált főnevek tanulásának hatékonysága nem tűnik az általános mentális rátermettséggel szorosan összefüggő képességnek, mégis közöltek olyan megfigyeléseket, amelyek szerint ez a teljesítmény is korrelál, bár lényegesen gyengébb kapcsolatot jelezve, az alvási orsók 2. stádiumú alvásra számított gyakoriságával (GAIS, MÖLLE és munkatársai, 2002), illetve az alvás első felének 3–4. stádiumaira számított intenzitásával (BERNER, SCHABUS és munkatársai, 2006). Ez látszólag ellentétben áll FOGEL, NADER és munkatársai (2007) fönt említett eredményével, akik a nem verbális képességterületeknek az alvási orsókkal való összefüggését találták, ugyanakkor fölvet egy másik szempontot is. Ez a feladatok megoldásakor alkalmazott egyéni kognitív stratégiák alkalmazásának kérdése, amit a memóriavizsgálatokban és az intelligenciatesztekben egyaránt nagyon nehéz kontrollálni. Vizuális-téri (non-verbális) képességterületen a Rey–Osterrieth Komplex Ábra Tesztben nyújtott felidézési teljesítmény és a 2. stádiumú alvásban a jobb parietális régió fölött mért gyors alvási orsók percenkénti gyakorisága között találtunk összefüggést (BÓDIZS, LÁZÁR, RIGÓ, 2008). Ez az eredmény, korábbi megfigyelésünkkel összevetve a kérgi lokalizáció alvási-orsózáson alapuló megközelítésének lehetőségével kecsegtet, hiszen az intelligencia neurobiológiáját a prefrontális kéreg strukturális és funkcionális jellemzőivel (GRAY, THOMPSON, 2004), míg a vizuális-téri emlékezeti teljesítményt a jobb agykéreg térfogatával (FJELL, WALHOVD és munkatársai, 2005) és a jobb parietális lebeny működésével (MOSCOVITCH, KAPUR és munkatársai, 1995) hozták összefüggésbe. Eredményeink ugyanakkor az intelligencia- és a frontális területek fölötti alvási orsózás (BÓDIZS, KIS és munkatársai, 2005), illetve a vizuális-téri emlékezet és a jobb parietális terület fölötti alvási orsózás (BÓDIZS, LÁZÁR, RIGÓ, 2008) kapcsolatát jelzik. Természetesen a két alvás-EEG mutató korántsem független egymástól, ami újra fölveti az intelligencia és a specifikus kognitív képességek kapcsolatának problémáját. A Rey–Osterrieth Komplex Ábra Teszt vonatkozásában ellentmondásos a szakirodalom, ugyanis alacsony korrelációról (FÖLDI, TOMASOVSZKY, 2003), illetve szorosabb összefüggésről (GALLAGHER, BURKE, 2007) egyaránt beszámoltak. Saját mintánkon az alacsony korreláció, tehát az intelligenciától való függetlenség bizonyult érvényesnek (BÓDIZS, LÁZÁR, RIGÓ, 2008). Az alvási orsózás neuropszichológiai lokalizáló szerepére utalnak még azok a kutatási eredmények is, amelyeket az alvás ideje alatti emlékezeti konszolidáció vizsgálata céljából végeztek. A bal frontális területek fölött regisztrált alvási orsók száma a verbális (CLEMENS, FABÓ, HALÁSZ, 2005), míg a jobb parietális területek fölötti alvási orsók száma a vizuális-téri (Rey–Osterrieth Komplex Ábra; CLEMENS, FABÓ, HALÁSZ, 2006) emlékezeti megőrzés hatékonyságával korrelál pozitívan. Bár, mint a fenti rövid összefoglalóból látszik, az alvási orsók és a kognitív teljesítmény közötti vonásszerű korrelatív kapcsolat korántsem tekinthető lezárt kutatási témának (1. táblázat), régről ismert és új eredmények is támogatják a terület kognitív neuropszichiátriai relevanciáját. Ezek közé a kutatási eredmények közé tartoznak a mentális retardációban (SHIBAGAKI, 2005), az időskori kognitív hanyatlásban és neurodegeneratív megbetegedésekben (PETIT, GAGNON és munkatársai, 2004), illetve az Aszperger-szindrómában (LIMOGES, MOTTRON és munkatársai, 2005) és a szkizofréniában (FERRARELLI, HUBER és munkatársai, 2007) észlelt módosult vagy csökkent orsózási mintázatok. Ezeknek a megfigyeléseknek a zöme az alapkutatás klinikai validitásvizsgálataként is felfogható. Így például csökkent alvási orsózás jellemző a Creutzfeldt–Jakob-kór korai stádiumára (DONNET, FARNARIER és munkatársai, 1992), az Alzheimer-kórra (PETIT, GAGNON és munkatársai, 2004) és a szkizofréniára (FERRARELLI, HUBER és munkatársai, 2007), miközben mindhárom rendellenesség kérgi atrófiával (FINKENSTAEDT, SZUDRA és munkatársai, 1996; KARAS, SCHELTENS és munkatársai, 2004; ANDREONE, TANSELLA és munkatársai, 2007) és többek között a vizuális-téri memória markáns zavarával jár (CHERRIER, MENDEZ és munkatársai, 1999; KAPUR, ABBOTT és munkatársai, 2003; GÖDER, BOIGS és munkatársai, 2004).
Állapotok és vonások: akut hatások leképeződése az alvás-EEG-benAz alvás-EEG egyénre jellemző volta, időbeli stabilitása és egyes kognitív teljesítményekkel való vonásszerű korrelációja nem jelenti azt, hogy az embert napközben ért hatások egyáltalán nem befolyásolják a kérdéses mutatókat. A lassú hullámú EEG-aktivitás alvás közben mért és topografikusan specifikus fokozódását lehet elérni olyan napközbeni hatásokkal, mint az egyik kart ért több órás szomatoszenzoros stimuláció (KATTLER, DIJK, BORBÉLY, 1994) vagy egyes vizuomotoros koordinációs feladatok (HUBER, GHILARDI és munkatársai, 2004). Utóbbi esetben a tanulásnak megfelelő agyi régió fölött a lassú hullámú EEG-ben mért lokális fokozódás pozitívan korrelált az éjszaka során, gyakorlás híján létrejött teljesítményfokozódással. Hasonló jelenségeket írtak le az alvási orsókkal kapcsolatban. Kimutatták, hogy az alvást megelőző deklaratív, verbális tanulás fokozza az alvási orsók gyakoriságát (GAIS, MÖLLE és munkatársai, 2002; SCHABUS, GRUBER és munkatársai, 2004). Igaz az orsófokozódás mértéke és az emlékezeti megőrzés hatékonysága között csak az egyik kutatásban találtak összefüggést (SCHABUS, GRUBER és munkatársai, 2004), de az orsózásukat a tanulás hatására fokozó személyek az alapszinten is erőteljesen orsózók közül kerültek ki. Így, ha az e téren ismert vonásszerű összefüggéseket is figyelembe vesszük, nem világos, hogy az orsók által biztosított hatékony emlékezeti megőrzésről vagy inkább a kognitívan hatékony és alapszinten is erőteljes orsózással jellemezhető személyek esetén hatékony kódolásról beszélhetünk (STICKGOLD, 2004). Motoros tanulást követően is leírták az alvási orsók megszaporodását (FOGEL, SMITH, 2006), ami elsősorban a jobb mozgástanulási készségekkel jellemezhető kísérleti személyekre jellemző (PETERS, SMITH, SMITH, 2007). A bal kézre szorítkozó mozgásos készségelsajátítás esetében az orsózás fokozódása, az elvártnak megfelelően a jobb centrális régióban mutatkozott meg (NISHIDA, WALKER, 2007). A fenti eredmények tehát kevés kétséget hagynak afelől, hogy a tanulás különböző formái kihatnak az alvásfüggő EEG-oszcillációkra. Az is több eredményből kiviláglik, hogy a személyek közötti különbségek befolyással vannak a tanulásindukált alvásmódosulásra (SCHABUS, GRUBER és munkatársai, 2004; PETERS, SMITH, SMITH, 2007). Jogosan merül tehát fel a kérdés: mennyire megbízhatóak a vonásszerű jegyek az alvás-EEG-ben, illetve mennyire megbízhatóak az állapotszerű változások? A vonás versus állapot jelleg közötti ellentmondás feloldása céljából eddig két vizsgálat született, átfedő mintákon. Az egyik egyértelműen rámutatott arra, hogy az alvási orsózás deklaratív és procedurális tanulás előtt és után egyaránt összefügg a Raven-féle Progresszív Mátrix Teszt eredményeivel, tehát elvileg a vonásszerűséget helyezte előtérbe (SCHABUS, HODLMOSER és munkatársai, 2006). A másik, némileg összetettebb elemzés nyomán kiderült, hogy az alvási orsók a deklaratív verbális tanulást követően növekedhetnek ugyan, és ez a növekedés kapcsolatban áll azzal, hogy a személyek mennyit őriznek meg a tanult anyagból, a változás léptéke azonban jóval kisebb a személyek közötti különbségeknél (SCHABUS, HOEDLMOSER és munkatársai, 2008). Vagyis mindkét hatás érvényes, de különböző skálákon kell mérni őket. Az egyének közötti különbségek nagyok és ezek a vonásszerű összefüggésekkel kapcsolatosak, míg az állapotszerű változások viszonylag kicsik, de szerepük jelentős, és amennyiben következetesen kimutathatóak, statisztikailag szignifikáns eredményekhez vezetnek. Egyéb alvás-EEG jegyek vonásszerű kognitív és affektív korrelátumai: az alfa hullámok problematikájaBár az alvás-EEG és a viselkedés, illetve a kogníció közötti kapcsolat vonásszerű aspektusait sokáig mostohán kezelte a szakirodalom, ennek ellenére elszórtan több olyan megfigyelés is született, amelyek valamilyen formában megfontolásra érdemesek, illetve közvetetten támogatják is az általunk fölvetett megközelítés érvényességét. Néhány ilyen jellegű adat az alvás alatt mért alfa (7–12 Hz) aktivitással kapcsolatos. Mivel az alfa hullámok leginkább a nyugodt, ingermentes ébrenlét jelei, alvás közbeni megjelenésükre sokáig úgy tekintettek, mint egyfajta megzavart, ébrenléthez közelebbi alvásállapotra, ami az álmatlansággal és a lassú hullámú alvással is kapcsolatos (SCHNEIDER-HELMERT, WHITEHOUSE és munkatársai, 2001). Ez a hipervigilencia-elmélet lényegében független a jelen felvetésben követett konnektivitás-plaszticitás megközelítéstől, de segíthet az olyan vonásszerű eredmények értelmezésében, mint amilyen a felnőttkori kötődés bizonytalansága és az alvás alatti alfa aktivitás túltengése közötti összefüggés (SLOAN, MAUNDER és munkatársai, 2007). A viselkedéses korrelátumok azonban nem minden esetben támasztják alá az egyszerű hipervigilencia-féle elképzelést. Azok az inszomniás páciensek, akiknek alvás közben sok alfa aktivitásuk van, szubjektív becslés alapján a valóságosnál hosszabb alvásról számolnak be, tehát nem érzik olyan rossznak az alvásukat, és ami még meglepőbb, egyben jobban teljesítenek a munkamemória számterjedelem próbájában is azoknál az inszomniásoknál, akikre a kevés alvás alatti alfa aktivitás jellemző (SCHNEIDER-HELMERT, KUMAR, 1995). Közismert, hogy a számterjedelem próbában nyújtott teljesítmény összefügg az intelligenciával (GRAY, THOMPSON, 2004), mint ahogy az is, hogy az intelligencia és az ébrenlét alatti alfa frekvencia, illetve gyors alfa komponens is megbízhatóan korrelálnak egymással (DOPPELMAYR, KLIMESCH és munkatársai, 2002). Az alvás alatti alfa aktivitás vonásszerű kognitív korrelátumainak kérdése ezért fontos felismerések lehetőségét rejti magában. Egy másik jól ismert összefüggés az ébrenlét alatti alfa hullámok frontális aszimmetriája és a pozitív, illetve negatív emocionalitás között áll fönn. Egy vizsgálatban kimutatták, hogy az ébrenlét állapotában mért alfa aszimmetria szendergésben (1. stádium) és REM alvásban is fönnmarad, vagyis az aktivált állapotok stabil és vonásszerű sajátosságának tekinthető. Ugyanakkor a 2. stádiumú alvásban mért alfa aszimmetria, meglepő módon fordítottan predikálta az emocionális diszpozíciókat: a jobb frontális alfa többlet (ami ébrenlétben csökkent frontális agyi aktivitásként értelmezhető) negatív emocionalitást jelzett előre (SCHMIDT, COTE és munkatársai, 2003). Ez az eredmény azonban értelmezhetővé válik, ha figyelembe vesszük az állapotok arousal-szintjeinek különbségeit: ami ébrenlétben inaktivitásnak minősül (alfa hullámok) a 2. stádiumú alvásban relatív aktivitástöbblet lehet. Vagyis a frontális alfa szimmetria és az emocionalitás kapcsolatát nemcsak ébrenlétben, de egyes alvásállapotokban is vizsgálhatjuk, figyelembe véve természetesen az adott alvásállapot sajátosságait. ÖSSZEGZÉSAz állapotok és vonások alvás-elektrofiziológiai markereinek kérdését egy analógiával az izomzathoz hasonlíthatjuk. Ha megmérjük egy ember izomtömegét, akkor abból bizonyos fokig kikövetkeztethető a fizikai erőnléte és teljesítőképessége is. A személy ebben a tekintetben elég nagy bizonyossággal más személyekhez hasonlítható. Ha arra vagyunk kíváncsiak, hogy egy adott erőpróbában gyakorlás hatására mennyit fejlődött, akkor a saját magához mért (helyi) izomtömeggyarapodás ennek a változásnak egy biológiai markere és egyben a próba kivitelezésének és a fejlődésnek a mechanizmusa is. Az ismételt változások azonban tartós vonásokká válhatnak, például valamilyen sporttevékenység hatására. Visszatérve az alvás-EEG-ben mérhető oszcillációkra elmondható, hogy az alvás-EEG mintázatok éppúgy eredményei a korábbi tanulásnak, mint formálói az új emléknyomoknak. Vagyis tükrözik az idegrendszeri konnektivitást, de egyben fokozzák is azt. Az előbbire utal az a megfigyelés miszerint a kérgi szinaptikus kapcsolatok erősségét fokozó, kísérletileg előidézett hosszú távú potencírozás facilitálja az orsók megjelenését (WERK, HARBOUR, CHAPMAN, 2005). Utóbbira példa az a már említett hatás, miszerint az orsók ideje alatti ingerlés tartósabb módosulást idéz elő a válaszkészségben, mint az orsóktól független ingerlés (TIMOFEEV, GRENIER és munkatársai, 2002). Előbbi a vonásszerű, utóbbi az állapothatásokkal kapcsolatos megközelítésnek felel meg. IRODALOMACHERMANN, P., BORBÉLY, A. A. (1997) Low-frequency (<1 Hz) oscillations in the human sleep electroencephalogram. Neuroscience, 81, 213–222. AMZICA, F., STERIADE, M. (1995) Disconnection of intracortical synaptic linkages disrupts synchronization of a slow oscillation. Journal of Neuroscience, 15, 4658–4677. ANDERSON, C., HORNE, J. A. (2003) Prefrontal cortex: links between low frequency delta EEG in sleep and neuropsychological performance in healthy, older people. Psychophysiology, 40, 349–357. ANDREONE, N., TANSELLA, M., CERINI, R., RAMBALDELLI, G., VERSACE, A., MARRELLA, G., PERLINI, C., DUSI, N., PELIZZA, L., BALESTRIERI, M., BARBUI, C., NOSE, M., GASPARINI, A., BRAMBILLA, P. (2007) Cerebral atrophy and white matter disruption in chronic schizophrenia. European Archives of Psychiatry and Clinical Neuroscience, 257, 3–11. AZUMI, K., SHIRAKAWA, S. (1982) Characteristics of spindle activity and their use in evaluation of hypnotics. Sleep, 5, 95–105. BERNER, I., SCHABUS, M., WIENERROITHER, T., KLIMESCH, W. (2006) The significance of sigma neurofeedback training on sleep spindles and aspects of declarative memory. Applied Psychophysiology and Biofeedback, 31, 97–114. BÓDIZS R. (2000) Alvás, álom, bioritmusok. Medicina könyvkiadó, Budapest BÓDIZS R. (2006) Megragadható-e a személyiség és az intellektus a frontális alvás-EEG finomelemzése által? In HALÁSZ P. (szerk.) Frontális lebeny mint a neurológia és a pszichiátria közös területe. 91–102. Melinda kiadó, Budapest BÓDIZS, R., BÉKÉSY, M., SZŰCS, A., BARSI, P., HALÁSZ, P. (2002) Sleep-dependent hippocampal slow activity correlates with waking memory performance in humans. Neurobiology of Learning and Memory, 78, 441–457. BÓDIZS, R., KIS, T., LÁZÁR, A. S., RIGÓ, P., HAVRÁN, L., CLEMENS, Z., HALÁSZ, P. (2005) Prediction of general mental ability based on neural oscillation measures of sleep. Journal of Sleep Research, 14, 285–292. BÓDIZS, R., LÁZÁR, A. S., RIGÓ, P. (2008) Correlation of visuospatial memory ability with right parietal EEG spindling during sleep. Acta Physiologica Hungarica (in press) BORBÉLY, A. A. (2001) From slow waves to sleep homeostasis: new perspectives. Archives Italiennes de Biologie, 139, 53–61. BUCKELMÜLLER, J., LANDOLT, H. P., STASSEN, H. H., ACHERMANN, P. (2006) Trait-like individual differences in the human sleep electroencephalogram. Neuroscience, 138, 351– 356. CAMPBELL, I. G., HIGGINS, L. M., DARCHIA, N., FEINBERG, I. (2006) Homeostatic behavior of fast Fourier transform power in very low frequency non-rapid eye movement human electroencephalogram. Neuroscience, 140, 1395–1399. CHERRIER, M. M., MENDEZ, M. F., DAVE, M., PERRYMAN, K. M. (1999) Performance on the Rey-Osterrieth Complex Figure Test in Alzheimer disease and vascular dementia. Neuropsychiatry, Neuropsychology and Behavioral Neurology, 12, 95–101. CLEMENS, Z., FABÓ, D., HALÁSZ, P. (2005) Overnight verbal memory retention correlates with the number of sleep spindles. Neuroscience, 132, 529–535. CLEMENS, Z., FABÓ, D., HALÁSZ, P. (2006) Twenty-four hours retention of visuospatial memory correlates with the number of parietal sleep spindles. Neuroscience Letters, 403, 52– 56. DAVIDSON, R. J. (2001) Toward a biology of personality and emotion. Annals of the New York Academy of Sciences, 935, 191–207. DE GENNARO, L., FERRARA, M. (2003) Sleep spindles: an overview. Sleep Medicine Reviews, 7, 423–440. DE GENNARO, L., FERRARA, M., VECCHIO, F., CURCIO, G., BERTINI, M. (2005) An electroencephalographic fingerprint of human sleep. Neuroimage, 26, 114–122. DONNET, G., FARNARIER, D., GAMBARELLI, U., AGUGLIA, H., REGIS, H. (1992) Sleep electroencephalogram at the early stage of Creutzfeldt-Jakob disease. Clinical Electroencephalography, 23, 118–125. DOPPELMAYR, M., KLIMESCH, W., StADLER, W., PÖLLHUBER, D., HEINE, C. (2002) EEG alpha power and intelligence. Intelligence, 30, 289–302. ESSER, S. K., HILL, S. L., TONONI, G. (2007) Sleep Homeostasis and Cortical Synchronization: I. Modeling the Effects of Synaptic Strength on Sleep Slow Waves. Sleep, 30, 1617– 1630. FERRARELLI, F., HUBER, R., PETERSON, M. J., MASSIMINI, M., MURPHY, M., RIEDNER, B. A., WATSON, A., BRIA, P., TONONI, G. (2007) Reduced sleep spindle activity in schizophrenia patients. American Journal of Psychiatry, 164, 483–492. FINELLI, L. A., ACHERMANN, P., BORBÉLY, A. A. (2001) Individual ‘fingerprints’ in human sleep EEG topography. Neuropsychopharmacology, 25, S57–S62. FINKENSTAEDT, M., SZUDRA, A., ZERR, I., POSER, S., HISE, J. H., STOEBNER, J. M., WEBER, T. (1996) MR imaging of Creutzfeldt-Jakob disease. Radiology, 199, 793–798. FJELL, A. M., WALHOVD, K. B., REINVANG, I., LUNDERVOLD, A., DALE, A. M., QUINN, B. T., MAKRIS, N., FISCHL, B. (2005) Age does not increase rate of forgetting over weeks – Neuroanatomical volumes and visual memory across the adult life-span. Journal of the International Neuropsychological Society, 11, 2–15. FOGEL, S. M., NADER, R., COTE, K. A., SMITH, C. T. (2007) Sleep spindles and learning potential. Behavioral Neuroscience, 121, 1–10. FOGEL, S. M., SMITH, C. T. (2006) Learning-dependent changes in sleep spindles and Stage 2 sleep. Journal of Sleep Research, 15, 250–255. FÖLDI R., TOMASOVSZKI L. (2003) A Rey-féle összetett figura és felismerési próba neuropszichológiai alkalmazási lehetőségei. Ideggyógyászati Szemle, 56, 82–91. GAILLARD, J. M., BLOIS, R. (1981) Spindle density in sleep of normal subjects. Sleep, 4, 385– 391. GAIS, S., MÖLLE, M., HELMS, K., BORN, J. (2002) Learning-dependent increases in sleep spindle density. Journal of Neuroscience, 22, 6830–6834. GALLAGHER, C., BURKE, T. (2007) Age, gender and IQ effects on the Rey-Osterrieth Complex Figure Test. British Journal of Clinical Psychology, 46 (Pt 1), 35–45. GÖDER, R., ALDENHOFF, J. B., BOIGS, M., BRAUN, S., KOCH, J., FRITZER, G. (2006) Delta power in sleep in relation to neuropsychological performance in healthy subjects and schizophrenia patients. Journal of Neuropsychiatry and Clinical Neurosciences, 18, 529–535. GÖDER, R., BOIGS, M., BRAUN, S., FRIEGE, L., FRITZER, G., ALDENHOFF, J. B., HINZE-SELCH, D. (2004) Impairment of visual memory is associated with decreased slow wave sleep in schizophrenia. Journal of Psychiatric Research, 38, 591–599. GRAY, J. R., THOMPSON, P. M. (2004) Neurobiology of intelligence: science and ethics. Nature Reviews Neuroscience, 5, 471–482. HUBER, R., GHILARDI, M. F., MASSIMINI, M., TONONI, G. (2004) Local sleep and learning. Nature, 430, 78–81. KAPUR, N., ABBOTT, P., LOWMAN, A., WILL, R. G. (2003) The neuropsychological profile associated with variant Creutzfeldt-Jakob disease. Brain, 126, 2693–2702. KARAS, G. B., SCHELTENS, P., ROMBOUTS, S. A., VISSER, P. J., VAN SCHIJNDEL, R. A., FOX, N. C., BARKHOF, F. (2004) Global and local gray matter loss in mild cognitive impairment and Alzheimer’s disease. Neuroimage, 23, 708–716. KATTLER, H., DIJK, D. J., BORBÉLY, A. A. (1994) Effect of unilateral somatosensory stimulation prior to sleep on the sleep EEG in humans. Journal of Sleep Research, 3 (3), 159–164. KIMURA, M., WINKELMANN, J. (2007) Genetics of sleep and sleep disorders. Cellular and Molecular Life Sciences, 64, 1216–1226. LIMOGES, E., MOTTRON, L., BOLDUC, C., BERTHIAUME, C., GODBOUT, R. (2005) Atypical sleep architecture and the autism phenotype. Brain, 128 (Pt 5), 1049–1061. LINKOWSKI, P., KERKHOFS, M., HAUSPIE, R., MENDLEWICZ, J. (1991) Genetic determinants of EEG sleep: a study in twins living apart. Electroencephalography and Clinical Neurophysiology, 79, 114–118. LINKOWSKI, P., KERKHOFS, M., HAUSPIE, R., SUSANNE, C., MENDLEWICZ, J. (1989) EEG sleep patterns in man: a twin study. Electroencephalography and Clinical Neurophysiology, 73, 279–284. LUBINSKI, D. (2000) Scientific and social significance of assessing individual differences: ‘‘sinking shafts at a few critical points’’. Annual Review of Psychology, 51, 405–444. MAGOUN, H. W. (1963) The waking brain. Charles C. Thomas Publisher, Springfield, Illinois MASSIMINI, M., FERRARELLI, F., ESSER, S. K., RIEDNER, B. A., HUBER, R., MURPHY, M., PETERSON, M. J., TONONI, G. (2007) Triggering sleep slow waves by transcranial magnetic stimulation. Proceedings of the National Academy of Sciences of the United States of America, 104, 8496–8501. MIYAMOTO, H., HENSCH, T. K. (2006) Bidirectional interaction of sleep and synaptic plasticity: A view from visual cortex. Sleep and Biological Rhythms, 4, 35–43. MIYAMOTO, H., KATAGIRI, H., HENSCH, T. (2003) Experience-dependent slow-wave sleep development. Nature Neuroscience, 6, 553–554. MÖLLE, M., MARSHALL, L., GAIS, S., BORN, J. (2002) Grouping of spindle activity during slow oscillations in human non-REM sleep. Journal of Neuroscience, 22, 10 941–10 947. MOSCOVITCH, C., KAPUR, S., KOHLER, S., HOULE, S. (1995) Distinct neural correlates of visual long-term memory for spatial location and object identity: a positron emission tomography study in humans. Proceedings of the National Academy of Sciences of the United States of America, 92, 3721–3725. NISHIDA, M., WALKER, M. P. (2007) Daytime naps, motor memory consolidation and regionally specific sleep spindles. PLoS ONE, 2 (4), e341. PETERS, K. R., SMITH, V., SMITH, C. T. (2007) Changes in sleep architecture following motor learning depend on initial skill level. Journal of Cognitive Neuroscience, 19, 817–829. PETIT, D., GAGNON, J. F., FANTINI, M. L., FERINI-STRAMBI, L. MONTPLAISIR, J. (2004) Sleep and quantitative EEG in neurodegenerative disorders. Journal of Psychosomatic Research, 56, 487–496. SCHABUS, M., DANG-VU, T. T., ALBOUY, G., BALTEAU, E., BOLY, M., CARRIER, J., DARSAUD, A., DEGUELDRE, C., DESSEILLES, M., GAIS, S., PHILLIPS, C., RAUCHS, G., SCHNAKERS, C., STERPENICH, V., VANDEWALLE, G., LUXEN, A., MAQUET, P. (2007) Hemodynamic cerebral correlates of sleep spindles during human non-rapid eye movement sleep. Proceedings of the National Academy of Sciences of the United States of America, 104, 13 164–13 169. SCHABUS, M., GRUBER, G., PARAPATICS, S., SAUTER, C., KLÖSCH, G., ANDERER, P., KLIMESCH, W., SALETU, B., ZEITLHOFER, J. (2004) Sleep spindles and their significance for declarative memory consolidation. Sleep, 27, 1479–1485. SCHABUS, M., HODLMOSER, K., GRUBER, G., SAUTER, C., ANDERER, P., KLOSCH, G., PARAPATICS, S., SALETU, B., KLIMESCH, W., ZEITLHOFER, J. (2006) Sleep spindle-related activity in the human EEG and its relation to general cognitive and learning abilities. European Journal of Neuroscience, 23, 1738–1746. SCHABUS, M., HOEDLMOSER, K., PECHERSTOFER, T., ANDERER, P., GRUBER, G., PARAPATICS, S., SAUTER, C., KLOESCH, G., KLIMESCH, W., SALETU, B., ZEITLHOFER, J. (2008) Interindividual sleep spindle differences and their relation to learning related enhancements. Brain Research, 1191, 127–135. SCHMIDT, L. A., COTE, K. A., SANTESSO, D. L., MILNER, C. E. (2003) Frontal electroencephalogram alpha asymmetry during sleep: stability and its relation to affective style. Emotion, 3, 401–407. SCHNEIDER-HELMERT, D., KUMAR, A. (1995) Sleep, its subjective perception, and daytime performance in insomniacs with a pattern of alpha sleep. Biological Psychiatry, 37, 99– 105. SCHNEIDER-HELMERT, D., WHITEHOUSE, I., KUMAR, A., LIJZENGA, C. (2001) Insomnia and alpha sleep in chronic non-organic pain as compared to primary insomnia. Neuropsychobiology, 43, 54–58. SHIBAGAKI, M. (2005) Delta and spindle components in the integrated EEG during nocturnal sleep for infants with developmental disabilities. Perceptual and Motor Skills, 101, 811–818. SILVERSTEIN, L. D., LEVY, C. M. (1976) The stability of the sigma sleep spindle. Electroencephalography and Clinical Neurophysiology, 40, 666–670. SLOAN, E. P., MAUNDER, R. G., HUNTER, J. J., MOLDOFSKY, H. (2007) Insecure attachment is associated with the alpha-EEG anomaly during sleep. BioPsychoSocial Medicine, 1, 20. STERIADE, M. (2003) The corticothalamic system is sleep. Frontiers in Biosciences, 8, d878– d899. STERIADE, M. (2006) Grouping of brain rhythms in corticothalamic systems. Neuroscience, 137, 1087–1106. STICKGOLD, R. (2004) Dissecting sleep-dependent learning and memory consolidation. Comment on Schabus M et al. Sleep spindles and their significance for declarative memory consolidation. Sleep, 27 (8), 1479–1485.; Sleep, 27, 1443–1445. TAN, X., CAMPBELL, I. G., FEINBERG, I. (2001) Internight reliability and benchmark values for computer analyses of non-rapid eye movement (NREM) and REM EEG in normal young adult and elderly subjects. Clinical Neurophysiology, 112, 1540–1552. TAN, X., CAMPBELL, I. G., PALAGINI, L., FEINBERG, I. (2000) High internight reliability of computer-measured NREM delta, sigma, and beta: biological implications. Biological Psychiatry, 48, 1010–1019. TIMOFEEV, I., GRENIER, F., BAZHENOV, M., HOUWELING, A. R., SEJNOWSKI, T. J., STERIADE, M. (2002) Short- and medium-term plasticity associated with augmenting responses in cortical slabs and spindles in intact cortex of cats in vivo. Journal of Physiology, 542 (Pt 2), 583–598. TUCKER, A. M., DINGES, D. F., VAN DONGEN, H. P. (2007) Trait interindividual differences in the sleep physiology of healthy young adults. Journal of Sleep Research, 16, 170–180. WERK, C. M., HARBOUR, V. L., CHAPMAN, C. A. (2005) Induction of long-term potentiation leads to increased reliability of evoked neocortical spindles in vivo. Neuroscience, 131, 793–800. WERTH, E., ACHERMANN, P., DIJK, D. J., BORBÉLY, A. A. (1997) Spindle frequency activity in the sleep EEG: individual differences and topographic distribution. Electroencephalography and Clinical Neurophysiology, 103, 535–542. |