
Essential Role of BMP4 Signaling in the Avian Ceca in Colorectal Enteric Nervous System Development
 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results
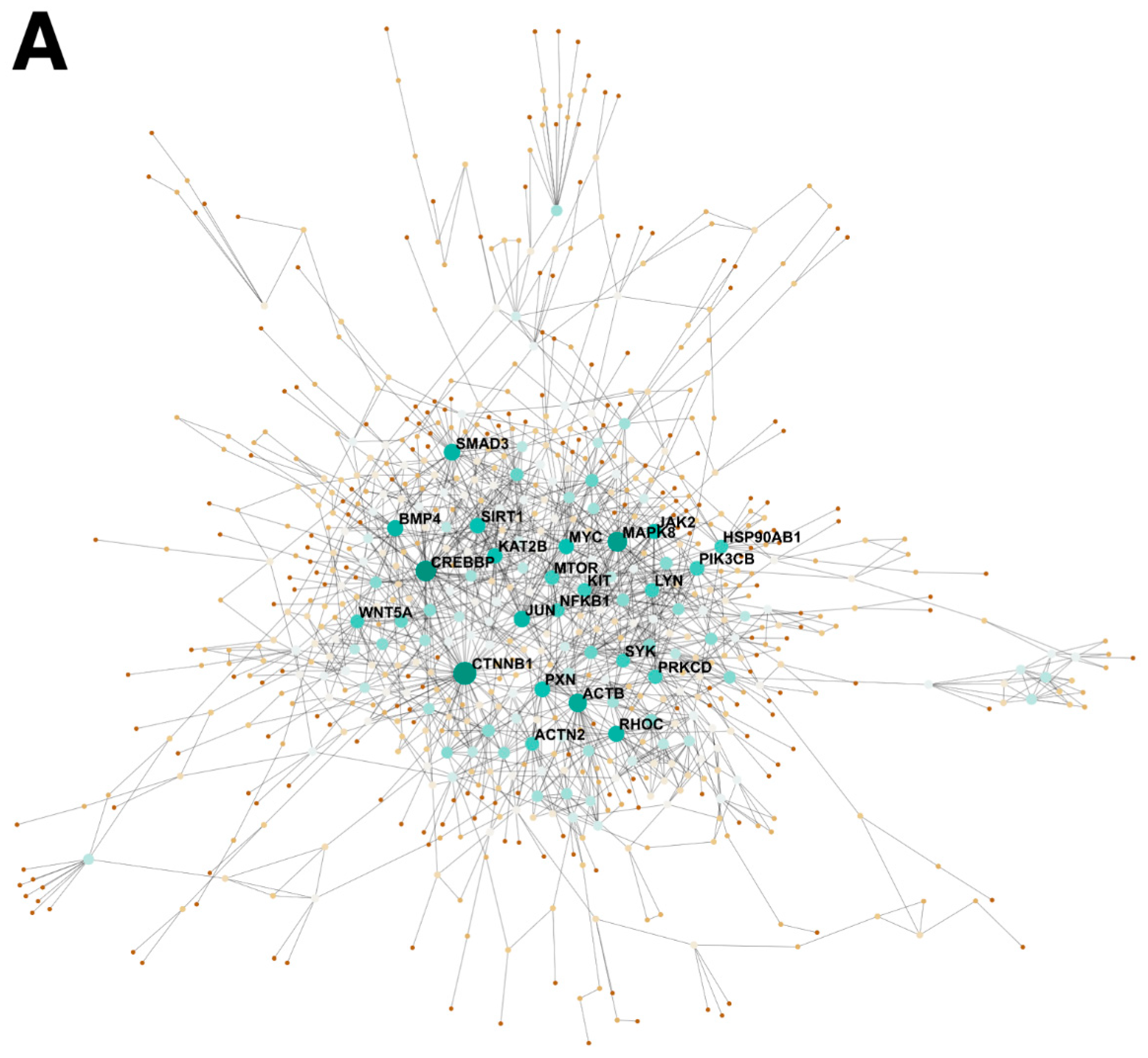
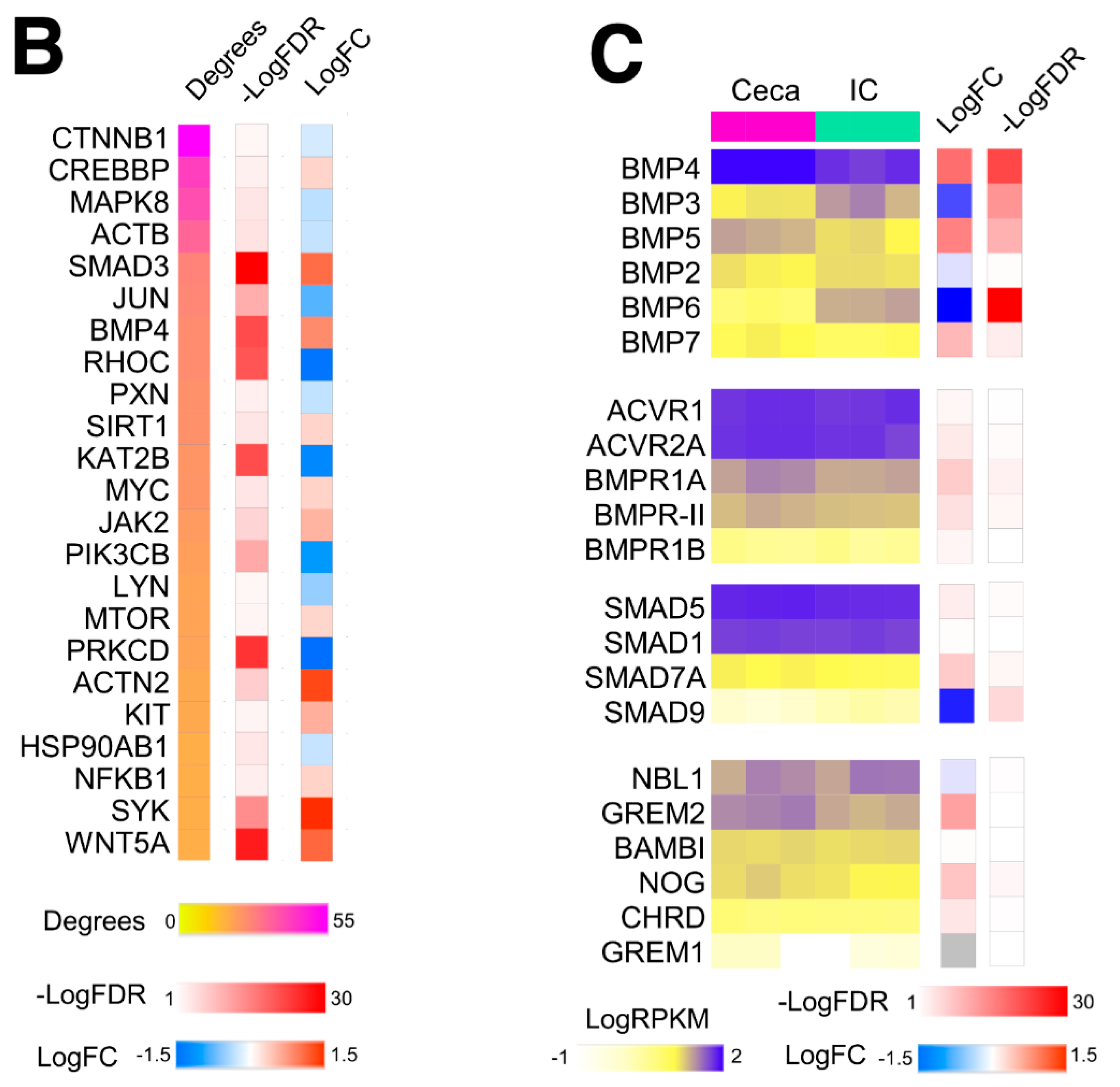
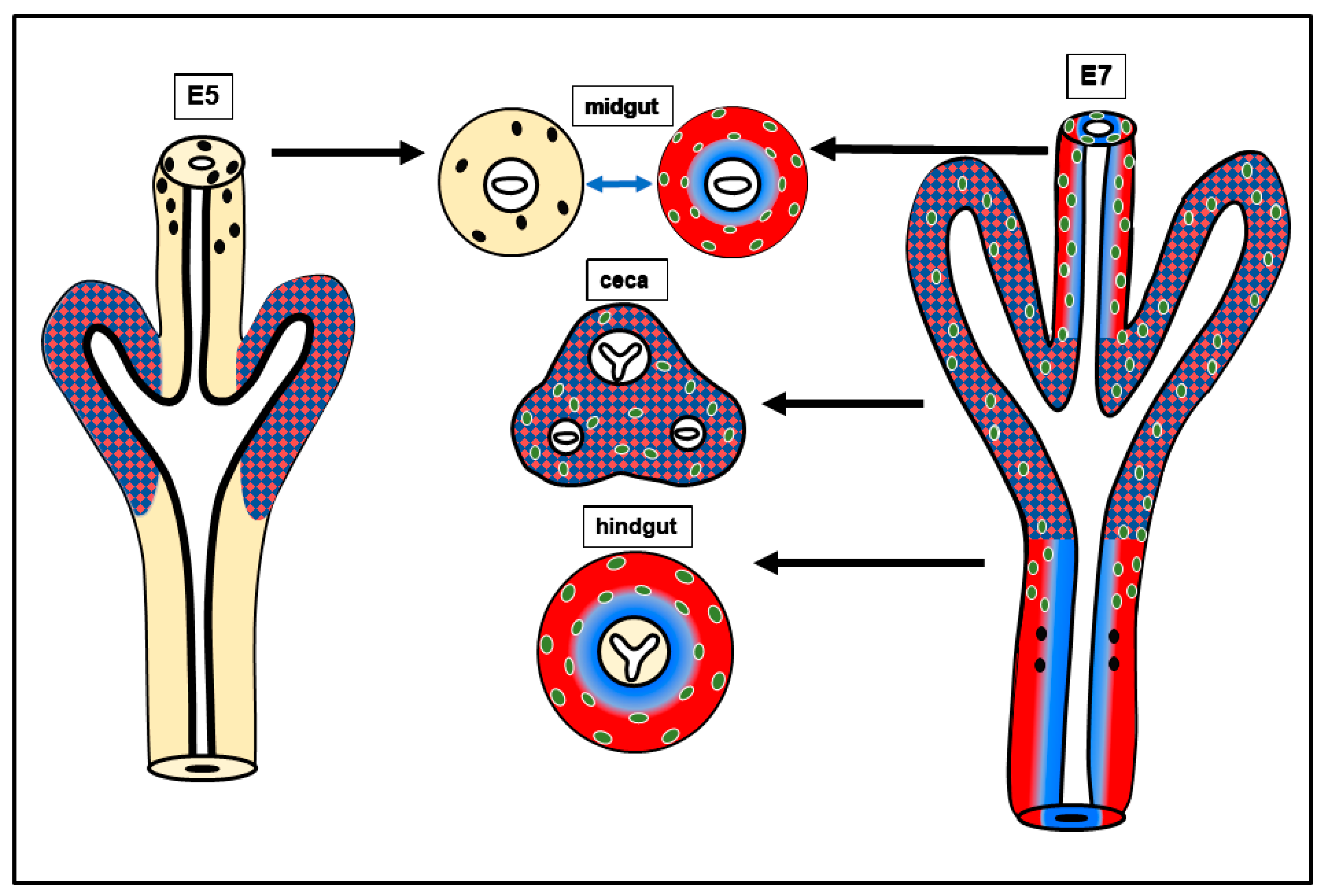
2.1. BMP4 Is An Important Signaling Hub in the Developing Intestine
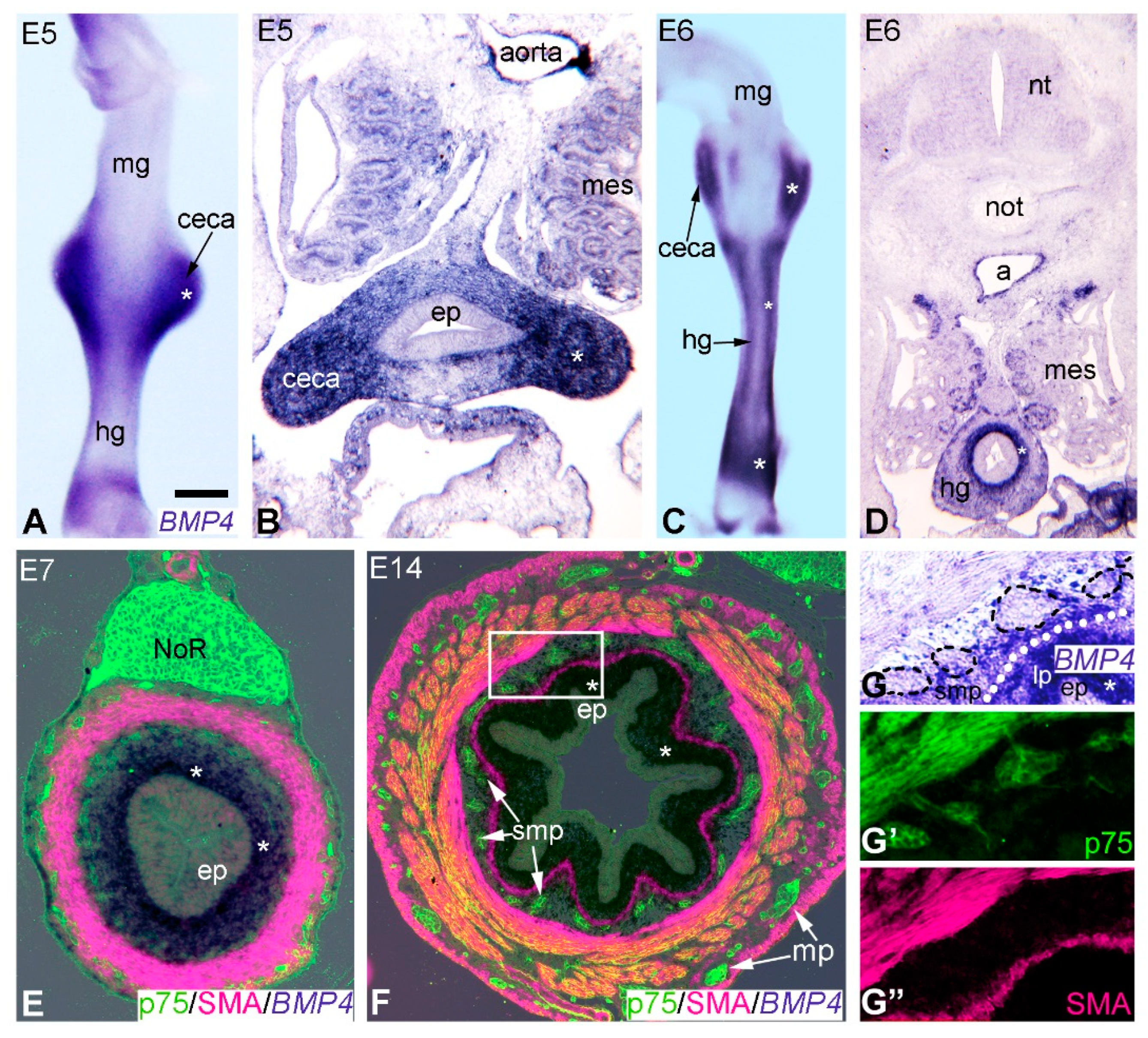
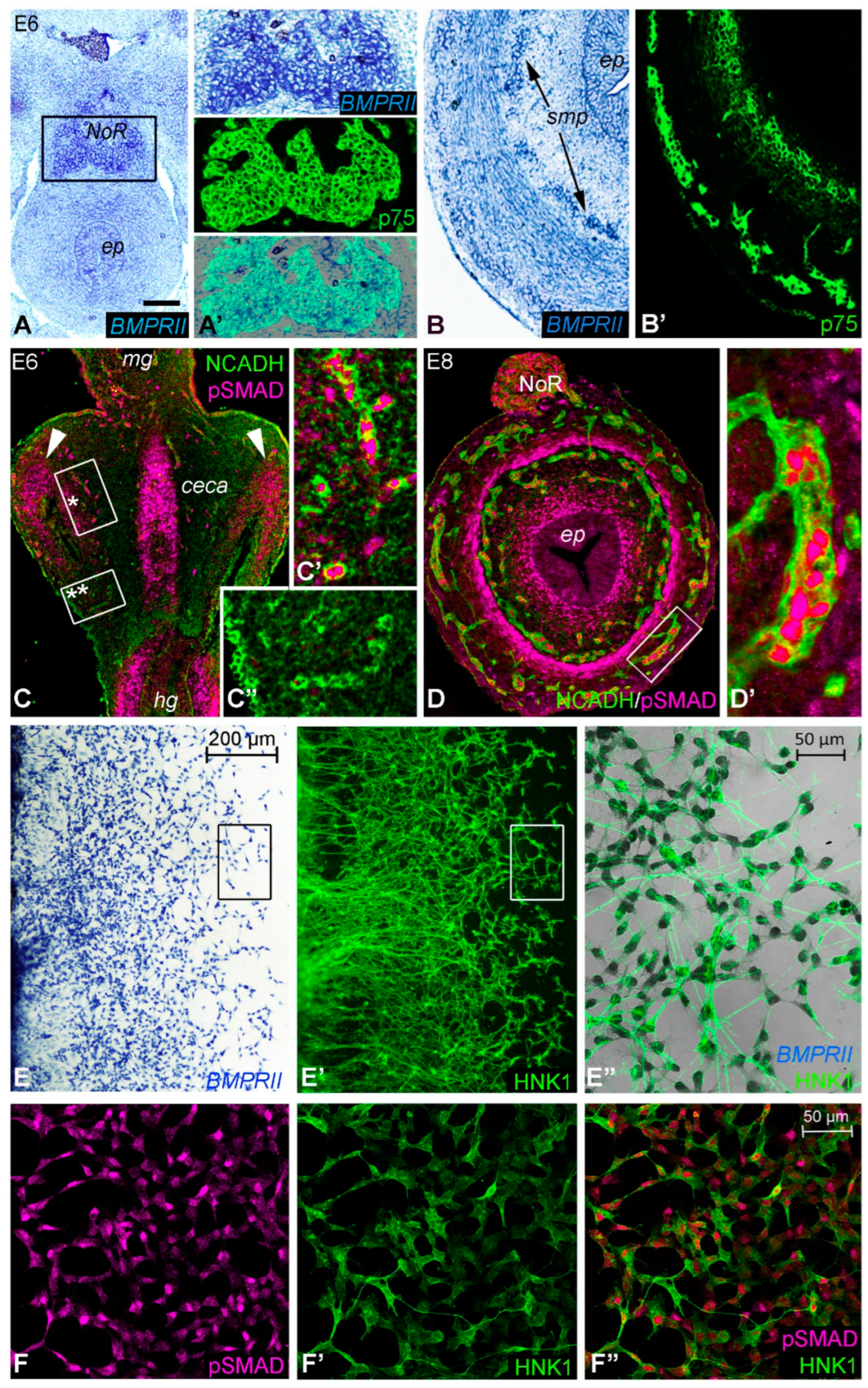
2.2. Expression of BMP4 Signaling Components Supports a Role during Hindgut ENS Formation
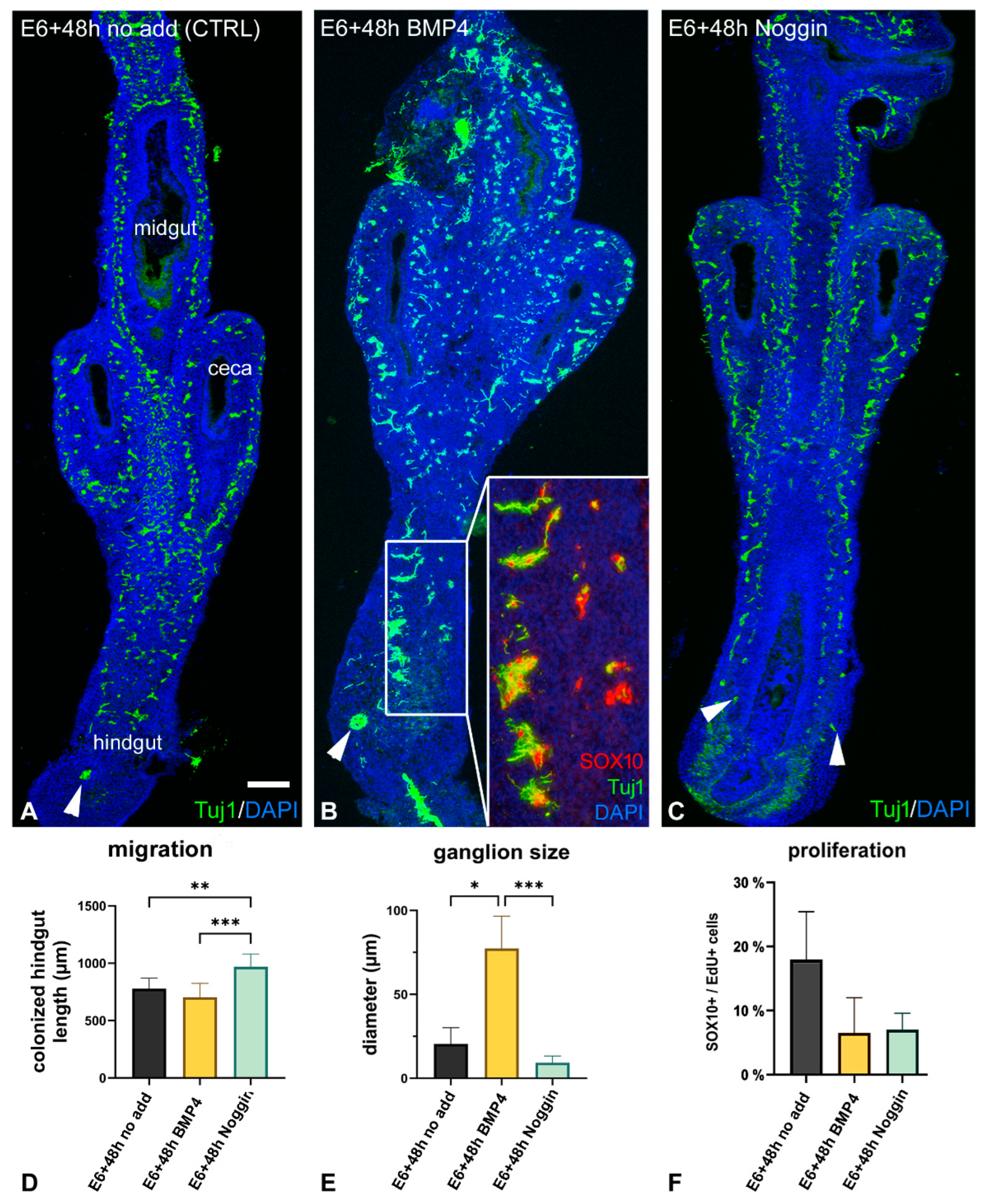
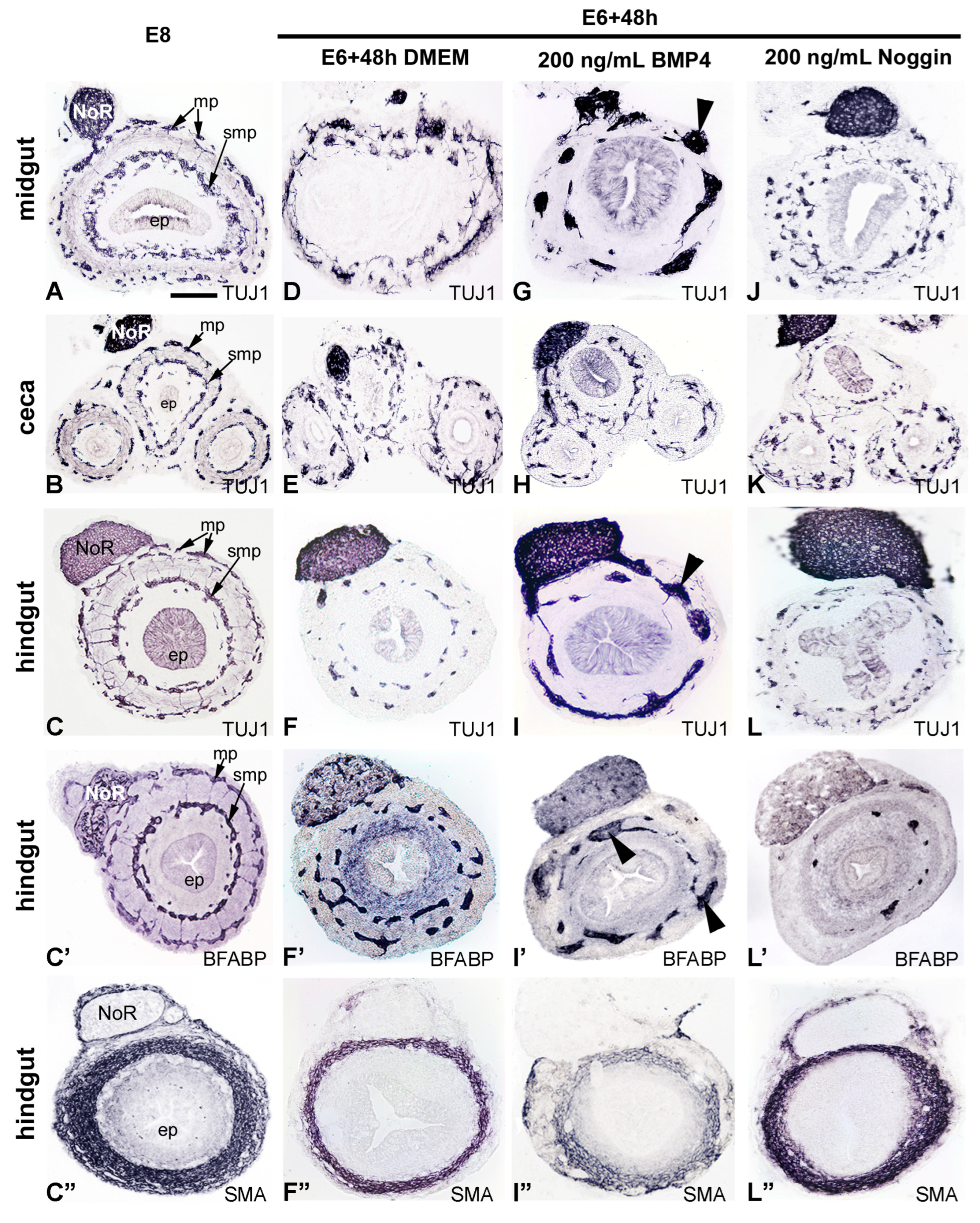
2.3. Inhibition of BMP4 Signaling Leads to Hindgut Hypoganglionosis
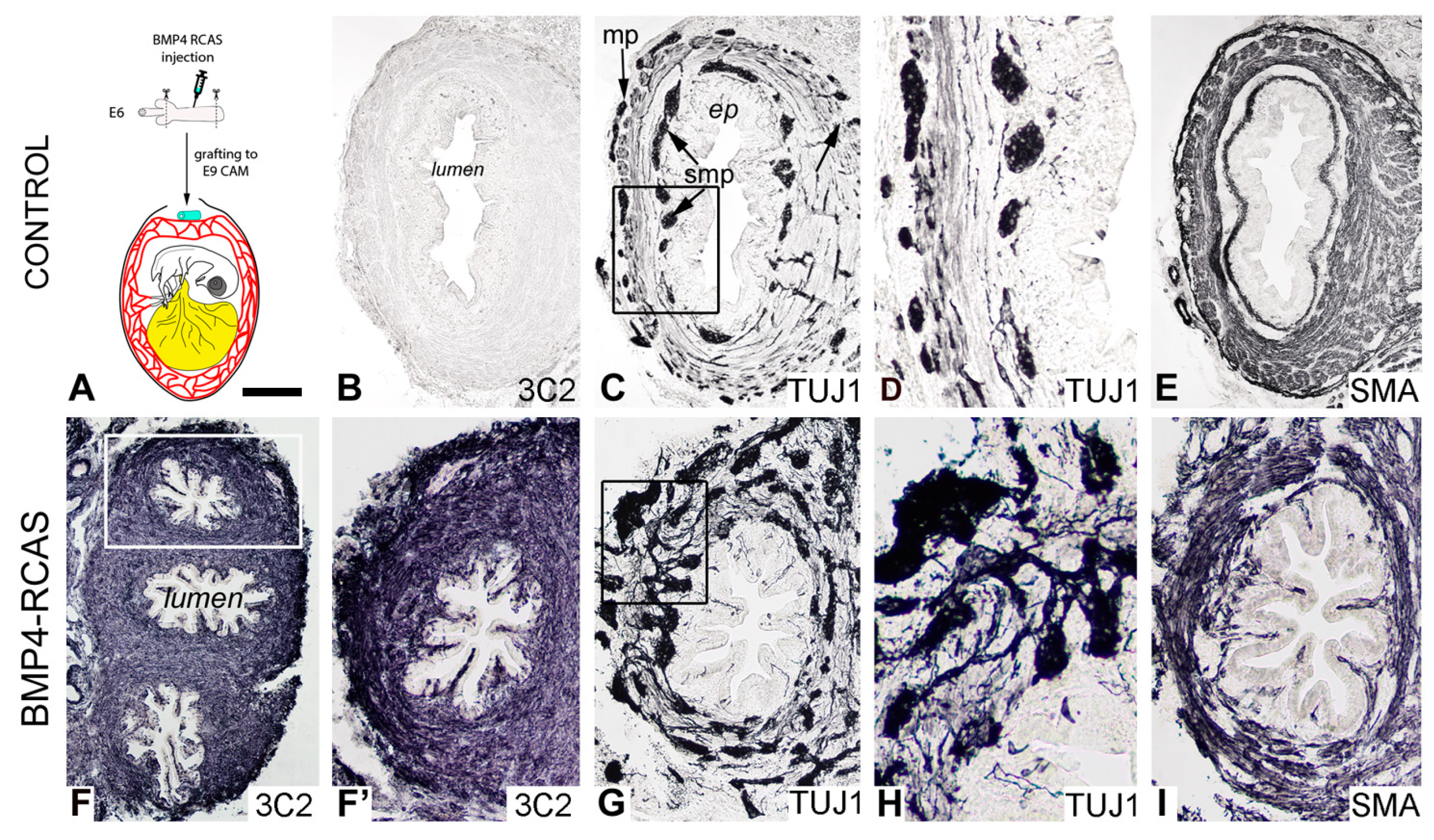
2.4. Retrovirus-Mediated Overexpression of BMP4 Induces Robust Gangliogenesis In Vivo
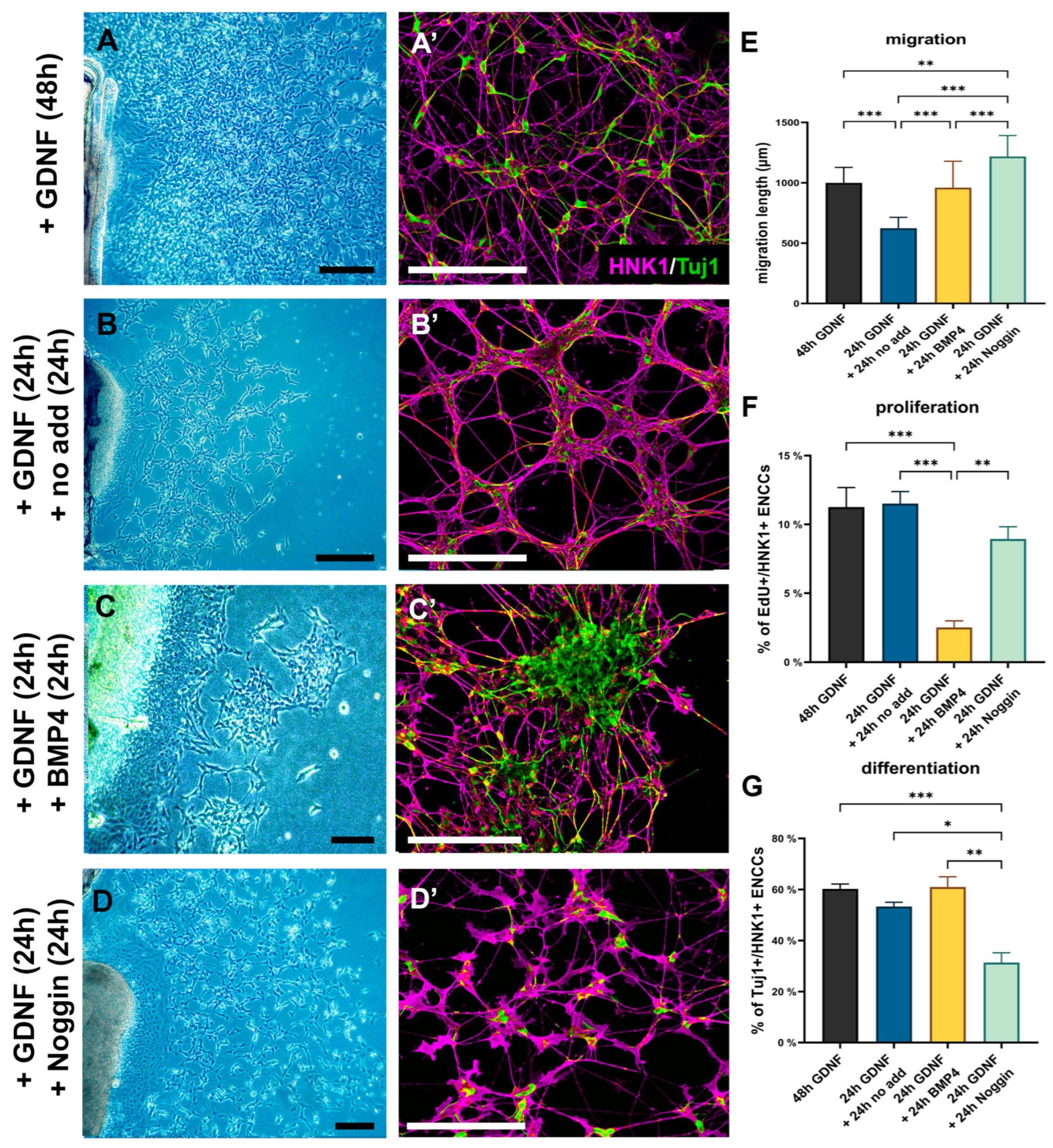
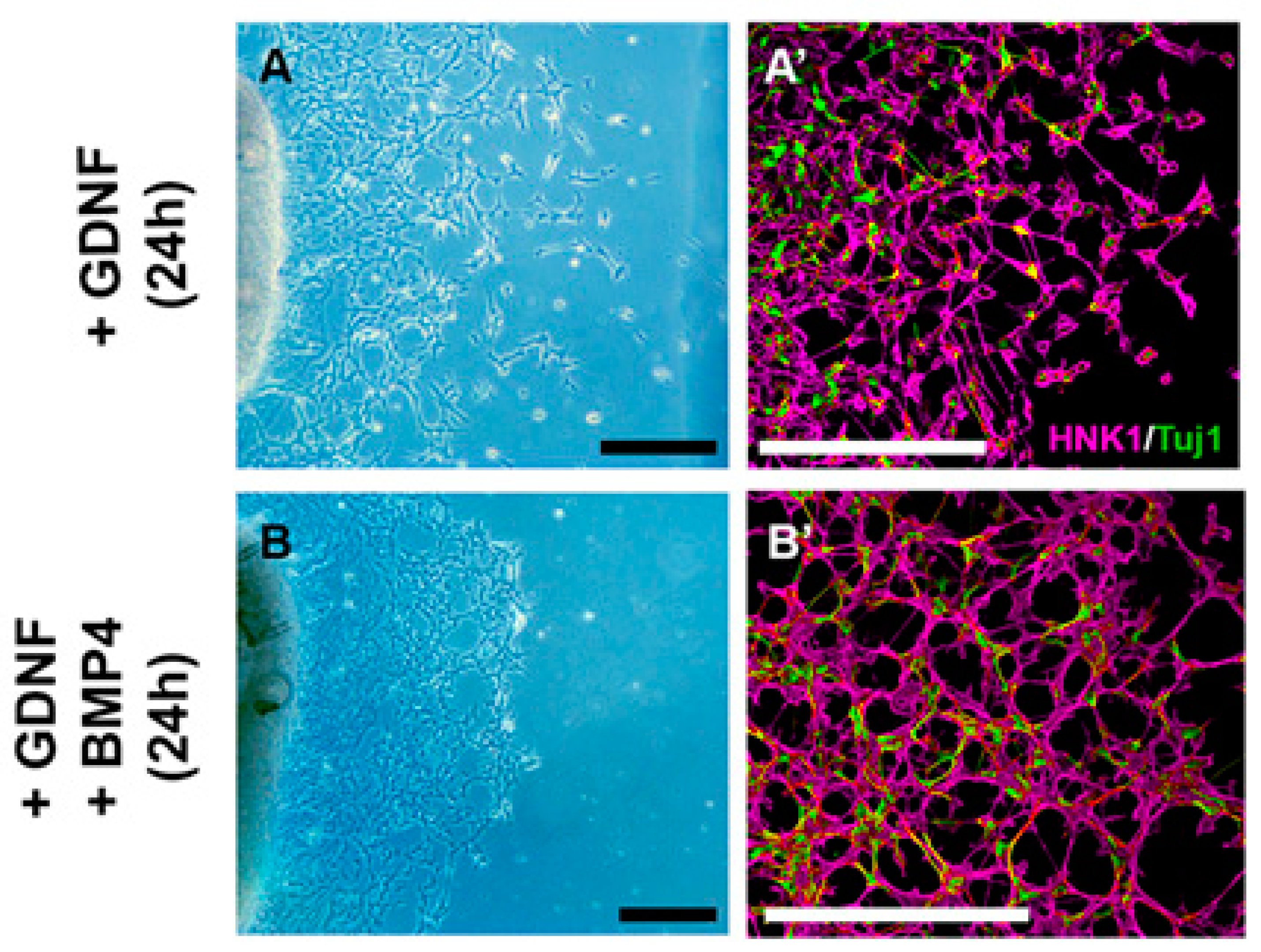
2.5. GDNF Inhibits BMP4-Induced ENCDC Aggregation
3. Discussion
4. Materials and Methods
4.1. Embryos
4.2. RNA-Seq
4.3. Immunohistochemistry
4.4. In Situ Hybridization on Sections and Whole-Mount
4.4.1. Whole-Mount In Situ Hybridization
4.4.2. In Situ Hybridization on FFPE Tissue Sections
4.5. Intestinal Organ Culture Assay
4.6. Viral Overexpression of BMP4 (+Chorioallantoic Membrane Transplantation)
4.7. Cell Migration Assay
4.8. EdU Labeling
4.9. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yntema, C.L.; Hammond, W.S. The Origin of Intrinsic Ganglia of Trunk Viscera from Vagal Neural Crest in the Chick Embryo. J. Comp. Neurol. 1954, 101, 515–541. [Google Scholar] [CrossRef]
- Burns, A.J.; Le Douarin, N.M. Enteric Nervous System Development: Analysis of the Selective Developmental Potentialities of Vagal and Sacral Neural Crest Cells Using Quail-Chick Chimeras. Anat. Rec. 2001, 262, 16–28. [Google Scholar] [CrossRef]
- Nagy, N.; Burns, A.J.; Goldstein, A.M. Immunophenotypic Characterization of Enteric Neural Crest Cells in the Developing Avian Colorectum. Dev. Dyn. 2012, 241, 842–851. [Google Scholar] [CrossRef]
- Catala, M.; Teillet, M.A.; Le Douarin, N.M. Organization and Development of the Tail Bud Analyzed with the Quail-Chick Chimaera System. Mech. Dev. 1995, 51, 51–65. [Google Scholar] [CrossRef]
- Burns, A.J.; Le Douarin, N.M. The Sacral Neural Crest Contributes Neurons and Glia to the Post- Umbilical Gut: Spatiotemporal Analysis of the Development of the Enteric Nervous System. Development 1998, 125, 4335–4347. [Google Scholar] [CrossRef]
- Nagy, N.; Brewer, K.C.; Mwizerwa, O.; Goldstein, A.M. Pelvic Plexus Contributes Ganglion Cells to the Hindgut Enteric Nervous System. Dev. Dyn. 2007, 236, 73–83. [Google Scholar] [CrossRef]
- Le Douarin, N.M.; Teillet, M.A. The Migration of Neural Crest Cells to the Wall of the Digestive Tract in Avian Embryo. J. Embryol. Exp. Morphol. 1973, 30, 31–48. [Google Scholar] [CrossRef]
- Kapur, R.P. Colonization of the Murine Hindgut by Sacral Crest-Derived Neural Precursors: Experimental Support for an Evolutionarily Conserved Model. Dev. Biol. 2000, 227, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Soret, R.; Schneider, S.; Bernas, G.; Christophers, B.; Souchkova, O.; Charrier, B.; Righini-Grunder, F.; Aspirot, A.; Landry, M.; Kembel, S.W.; et al. Glial Cell-Derived Neurotrophic Factor Induces Enteric Neurogenesis and Improves Colon Structure and Function in Mouse Models of Hirschsprung Disease. Gastroenterology 2020, 159, 1824–1838.e17. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, A.M.; Hofstra, R.M.W.; Burns, A.J. Building a Brain in the Gut: Development of the Enteric Nervous System. Clin. Genet. 2013, 83, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Uesaka, T.; Nagashimada, M.; Enomoto, H. Neuronal Differentiation in Schwann Cell Lineage Underlies Postnatal Neurogenesis in the Enteric Nervous System. J. Neurosci. 2015, 35, 9879–9888. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, D.J.; Bethell, G.S.; Shukla, R.; Kenny, S.E.; Edgar, D.H. Isolation of Enteric Nervous System Progenitor Cells from the Aganglionic Gut of Patients with Hirschsprung’s Disease. PLoS ONE 2015, 10, e0125724. [Google Scholar] [CrossRef] [PubMed]
- Pan, W.; Rahman, A.A.; Stavely, R.; Bhave, S.; Guyer, R.; Omer, M.; Picard, N.; Goldstein, A.M.; Hotta, R. Schwann Cells in the Aganglionic Colon of Hirschsprung Disease Can Generate Neurons for Regenerative Therapy. Stem Cells Transl. Med. 2022, 11, 1232–1244. [Google Scholar] [CrossRef]
- Lefèvre, M.A.; Soret, R.; Pilon, N. Harnessing the Power of Enteric Glial Cells’ Plasticity and Multipotency for Advancing Regenerative Medicine. Int. J. Mol. Sci. 2023, 24, 12475. [Google Scholar] [CrossRef] [PubMed]
- Uesaka, T.; Okamoto, M.; Nagashimada, M.; Tsuda, Y.; Kihara, M.; Kiyonari, H.; Enomoto, H. Enhanced Enteric Neurogenesis by Schwann Cell Precursors in Mouse Models of Hirschsprung Disease. Glia 2021, 69, 2575–2590. [Google Scholar] [CrossRef] [PubMed]
- Roberts, D.J. Molecular Mechanisms of Development of the Gastrointestinal Tract. Dev. Dyn. 2000, 219, 109–120. [Google Scholar] [CrossRef]
- Jiang, Y.; Liu, M.; Gershon, M.D. Netrins and DCC in the Guidance of Migrating Neural Crest-Derived Cells in the Developing Bowel and Pancreas. Dev. Biol. 2003, 258, 364–384. [Google Scholar] [CrossRef]
- Nagy, N.; Barad, C.; Graham, H.K.; Hotta, R.; Cheng, L.S.; Fejszak, N.; Goldstein, A.M. Sonic Hedgehog Controls Enteric Nervous System Development by Patterning the Extracellular Matrix. Development 2016, 143, 264–275. [Google Scholar] [CrossRef]
- Roberts, D.J.; Johnson, R.L.; Burke, A.C.; Nelson, C.E.; Morgan, B.A.; Tabin, C. Sonic Hedgehog Is an Endodermal Signal Inducing Bmp-4 and Hox Genes during Induction and Regionalization of the Chick Hindgut. Development 1995, 121, 3163–3174. [Google Scholar] [CrossRef]
- Roberts, D.J.; Smith, D.M.; Goff, D.J.; Tabin, C.J. Epithelial-Mesenchymal Signaling during the Regionalization of the Chick Gut. Development 1998, 125, 2791–2801. [Google Scholar] [CrossRef]
- Chalazonitis, A.; Kessler, J.A. Pleiotropic Effects of the Bone Morphogenetic Proteins on Development of the Enteric Nervous System. Dev. Neurobiol. 2012, 72, 843–856. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Stappenbeck, T.S.; White, A.C.; Lavine, K.J.; Gordon, J.I.; Ornitz, D.M. Reciprocal Epithelial-Mesenchymal FGF Signaling Is Required for Cecal Development. Development 2006, 133, 173–180. [Google Scholar] [CrossRef]
- Kruger, G.M.; Mosher, J.T.; Tsai, Y.-H.; Yeager, K.J.; Iwashita, T.; Gariepy, C.E.; Morrison, S.J. Temporally Distinct Requirements for Endothelin Receptor B in the Generation and Migration of Gut Neural Crest Stem Cells. Neuron 2003, 40, 917–929. [Google Scholar] [CrossRef]
- Nagy, N.; Kovacs, T.; Stavely, R.; Halasy, V.; Soos, A.; Szocs, E.; Hotta, R.; Graham, H.; Goldstein, A.M. Avian Ceca Are Indispensable for Hindgut Enteric Nervous System Development. Development 2021, 148, dev199825. [Google Scholar] [CrossRef]
- Fu, M.; Vohra, B.P.S.; Wind, D.; Heuckeroth, R.O. BMP Signaling Regulates Murine Enteric Nervous System Precursor Migration, Neurite Fasciculation, and Patterning via Altered Ncam1 Polysialic Acid Addition. Dev. Biol. 2006, 299, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, A.M.; Brewer, K.C.; Doyle, A.M.; Nagy, N.; Roberts, D.J. BMP Signaling Is Necessary for Neural Crest Cell Migration and Ganglion Formation in the Enteric Nervous System. Mech. Dev. 2005, 122, 821–833. [Google Scholar] [CrossRef] [PubMed]
- Chalazonitis, A.; Pham, T.D.; Li, Z.; Roman, D.; Guha, U.; Gomes, W.; Kan, L.; Kessler, J.A.; Gershon, M.D. Bone Morphogenetic Protein Regulation of Enteric Neuronal Phenotypic Diversity: Relationship to Timing of Cell Cycle Exit. J. Comp. Neurol. 2008, 509, 474–492. [Google Scholar] [CrossRef]
- Chalazonitis, A.; D’Autreaux, F.; Pham, T.D.; Kessler, J.A.; Gershon, M.D. Bone Morphogenetic Proteins Regulate Enteric Gliogenesis by Modulating ErbB3 Signaling. Dev. Biol. 2011, 350, 64–79. [Google Scholar] [CrossRef]
- Huycke, T.R.; Miller, B.M.; Gill, H.K.; Nerurkar, N.L.; Sprinzak, D.; Mahadevan, L.; Tabin, C.J. Genetic and Mechanical Regulation of Intestinal Smooth Muscle Development. Cell 2019, 179, 90–105.e21. [Google Scholar] [CrossRef]
- De Santa Barbara, P.; Williams, J.; Goldstein, A.M.; Doyle, A.M.; Nielsen, C.; Winfield, S.; Faure, S.; Roberts, D.J. Bone Morphogenetic Protein Signaling Pathway Plays Multiple Roles during Gastrointestinal Tract Development. Dev. Dyn. 2005, 234, 312–322. [Google Scholar] [CrossRef]
- Faure, C.; Chalazonitis, A.; Rheaume, C.; Bouchard, G.; Sampathkumar, S.G.; Yarema, K.J.; Gershon, M.D. Gangliogenesis in the Enteric Nervous System: Roles of the Polysialylation of the Neural Cell Adhesion Molecule and Its Regulation by Bone Morphogenetic Protein-4. Dev. Dyn. 2007, 236, 44–59. [Google Scholar] [CrossRef]
- Chalazonitis, A.; D’Autréaux, F.; Guha, U.; Pham, T.D.; Faure, C.; Chen, J.J.; Roman, D.; Kan, L.; Rothman, T.P.; Kessler, J.A.; et al. Bone Morphogenetic Protein-2 and -4 Limit the Number of Enteric Neurons but Promote Development of a TrkC-Expressing Neurotrophin-3-Dependent Subset. J. Neurosci. 2004, 24, 4266–4282. [Google Scholar] [CrossRef] [PubMed]
- Sukegawa, A.; Narita, T.; Kameda, T.; Saitoh, K.; Nohno, T.; Iba, H.; Yasugi, S.; Fukuda, K. The Concentric Structure of the Developing Gut Is Regulated by Sonic Hedgehog Derived from Endodermal Epithelium. Development 2000, 127, 1971–1980. [Google Scholar] [CrossRef] [PubMed]
- Pisano, J.M.; Colón-Hastings, F.; Birren, S.J. Postmigratory Enteric and Sympathetic Neural Precursors Share Common, Developmentally Regulated, Responses to BMP2. Dev. Biol. 2000, 227, 1–11. [Google Scholar] [CrossRef]
- Thang, S.H.; Kobayashi, M.; Matsuoka, I. Regulation of Glial Cell Line-Derived Neurotrophic Factor Responsiveness in Developing Rat Sympathetic Neurons by Retinoic Acid and Bone Morphogenetic Protein-2. J. Neurosci. 2000, 20, 2917–2925. [Google Scholar] [CrossRef] [PubMed]
- Ishizuya-Oka, A.; Hasebe, T. Sonic Hedgehog and Bone Morphogenetic Protein-4 Signaling Pathway Involved in Epithelial Cell Renewal along the Radial Axis of the Intestine. Digestion 2008, 77 (Suppl. S1), 42–47. [Google Scholar] [CrossRef]
- Amiel, J.; Lyonnet, S. Hirschsprung Disease, Associated Syndromes, and Genetics: A Review. J. Med. Genet. 2001, 38, 729–739. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Que, J. BMP Signaling in Development, Stem Cells, and Diseases of the Gastrointestinal Tract. Annu. Rev. Physiol. 2020, 82, 251–273. [Google Scholar] [CrossRef]
- van Grunsven, L.A.; Taelman, V.; Michiels, C.; Verstappen, G.; Souopgui, J.; Nichane, M.; Moens, E.; Opdecamp, K.; Vanhomwegen, J.; Kricha, S.; et al. XSip1 Neuralizing Activity Involves the Co-Repressor CtBP and Occurs through BMP Dependent and Independent Mechanisms. Dev. Biol. 2007, 306, 34–49. [Google Scholar] [CrossRef]
- Wei, W.; Liu, B.; Jiang, H.; Jin, K.; Xiang, M. Requirement of the Mowat-Wilson Syndrome Gene Zeb2 in the Differentiation and Maintenance of Non-Photoreceptor Cell Types During Retinal Development. Mol. Neurobiol. 2019, 56, 1719–1736. [Google Scholar] [CrossRef]
- Wakamatsu, N.; Yamada, Y.; Yamada, K.; Ono, T.; Nomura, N.; Taniguchi, H.; Kitoh, H.; Mutoh, N.; Yamanaka, T.; Mushiake, K.; et al. Mutations in SIP1, Encoding Smad Interacting Protein-1, Cause a Form of Hirschsprung Disease. Nat. Genet. 2001, 27, 369–370. [Google Scholar] [CrossRef] [PubMed]
- Cacheux, V.; Dastot-Le Moal, F.; Kääriäinen, H.; Bondurand, N.; Rintala, R.; Boissier, B.; Wilson, M.; Mowat, D.; Goossens, M. Loss-of-Function Mutations in SIP1 Smad Interacting Protein 1 Result in a Syndromic Hirschsprung Disease. Hum. Mol. Genet. 2001, 10, 1503–1510. [Google Scholar] [CrossRef] [PubMed]
- Nagaya, M.; Kato, J.; Niimi, N.; Tanaka, S.; Wakamatsu, N. Clinical Features of a Form of Hirschsprung’s Disease Caused by a Novel Genetic Abnormality. J. Pediatr. Surg. 2002, 37, 1117–1122. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.M.; Nielsen, C.; Tabin, C.J.; Roberts, D.J. Roles of BMP Signaling and Nkx2.5 in Patterning at the Chick Midgut-Foregut Boundary. Development 2000, 127, 3671–3681. [Google Scholar] [CrossRef]
- Shyer, A.E.; Huycke, T.R.; Lee, C.; Mahadevan, L.; Tabin, C.J. Bending Gradients: How the Intestinal Stem Cell Gets Its Home. Cell 2015, 161, 569–580. [Google Scholar] [CrossRef] [PubMed]
- Nerurkar, N.L.; Mahadevan, L.; Tabin, C.J. BMP Signaling Controls Buckling Forces to Modulate Looping Morphogenesis of the Gut. Proc. Natl. Acad. Sci. USA 2017, 114, 2277–2282. [Google Scholar] [CrossRef]
- Druckenbrod, N.R.; Epstein, M.L. Age-Dependent Changes in the Gut Environment Restrict the Invasion of the Hindgut by Enteric Neural Progenitors. Development 2009, 136, 3195–3203. [Google Scholar] [CrossRef]
- Wu, J.J.; Chen, J.X.; Rothman, T.P.; Gershon, M.D. Inhibition of in Vitro Enteric Neuronal Development by Endothelin-3: Mediation by Endothelin B Receptors. Development 1999, 126, 1161–1173. [Google Scholar] [CrossRef]
- Bondurand, N.; Natarajan, D.; Barlow, A.; Thapar, N.; Pachnis, V. Maintenance of Mammalian Enteric Nervous System Progenitors by SOX10 and Endothelin 3 Signalling. Development 2006, 133, 2075–2086. [Google Scholar] [CrossRef]
- Bondurand, N.; Dufour, S.; Pingault, V. News from the Endothelin-3/EDNRB Signaling Pathway: Role during Enteric Nervous System Development and Involvement in Neural Crest-Associated Disorders. Dev. Biol. 2018, 444 (Suppl. S1), S156–S169. [Google Scholar] [CrossRef]
- Mwizerwa, O.; Das, P.; Nagy, N.; Akbareian, S.E.; Mably, J.D.; Goldstein, A.M. Gdnf Is Mitogenic, Neurotrophic, and Chemoattractive to Enteric Neural Crest Cells in the Embryonic Colon. Dev. Dyn. 2011, 240, 1402–1411. [Google Scholar] [CrossRef]
- Sela-Donenfeld, D.; Kalcheim, C. Regulation of the Onset of Neural Crest Migration by Coordinated Activity of BMP4 and Noggin in the Dorsal Neural Tube. Development 1999, 126, 4749–4762. [Google Scholar] [CrossRef] [PubMed]
- Fu, M.; Tam, P.K.H.; Sham, M.H.; Lui, V.C.H. Embryonic Development of the Ganglion Plexuses and the Concentric Layer Structure of Human Gut: A Topographical Study. Anat. Embryol. 2004, 208, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Wang, Y.; Luo, L.; Li, X.; Jin, X.; Li, S.; Yu, X.; Yang, M.; Guo, Z. BMP2 Is Related to Hirschsprung’s Disease and Required for Enteric Nervous System Development. Front. Cell. Neurosci. 2019, 13, 523. [Google Scholar] [CrossRef] [PubMed]
- Anitha, M.; Shahnavaz, N.; Qayed, E.; Joseph, I.; Gossrau, G.; Mwangi, S.; Sitaraman, S.V.; Greene, J.G.; Srinivasan, S. BMP2 Promotes Differentiation of Nitrergic and Catecholaminergic Enteric Neurons through a Smad1-Dependent Pathway. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 298, G375–G383. [Google Scholar] [CrossRef] [PubMed]
- Hamburger, V.; Hamilton, H.L. A Series of Normal Stages in the Development of the Chick Embryo. J. Morphol. 1951, 88, 49–92. [Google Scholar] [CrossRef]
- Southwell, B.R. Staging of Intestinal Development in the Chick Embryo. Anat. Rec. A. Discov. Mol. Cell. Evol. Biol. 2006, 288A, 909–920. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor Package for Differential Expression Analysis of Digital Gene Expression Data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Zhou, G.; Soufan, O.; Ewald, J.; Hancock, R.E.W.; Basu, N.; Xia, J. NetworkAnalyst 3.0: A Visual Analytics Platform for Comprehensive Gene Expression Profiling and Meta-Analysis. Nucleic Acids Res. 2019, 47, W234–W241. [Google Scholar] [CrossRef]
- Moniot, B.; Biau, S.; Faure, S.; Nielsen, C.M.; Berta, P.; Roberts, D.J.; de Santa Barbara, P. SOX9 Specifies the Pyloric Sphincter Epithelium through Mesenchymal-Epithelial Signals. Development 2004, 131, 3795–3804. [Google Scholar] [CrossRef]
- Le Guen, L.; Notarnicola, C.; de Santa Barbara, P. Intermuscular Tendons Are Essential for the Development of Vertebrate Stomach. Dev. Camb. Engl. 2009, 136, 791–801. [Google Scholar] [CrossRef]
- Zou, H.; Niswander, L. Requirement for BMP Signaling in Interdigital Apoptosis and Scale Formation. Science 1996, 272, 738–741. [Google Scholar] [CrossRef]
- Jowett, J.B.; Xie, Y.; Chen, I.S.Y. The Presence of Human Immunodeficiency Virus Type 1 Vpr Correlates with a Decrease in the Frequency of Mutations in a Plasmid Shuttle Vector. J. Virol. 1999, 73, 7132–7137. [Google Scholar] [CrossRef]
- Riddle, R.D.; Johnson, R.L.; Laufer, E.; Tabin, C. Sonic Hedgehog Mediates the Polarizing Activity of the ZPA. Cell 1993, 75, 1401–1416. [Google Scholar] [CrossRef] [PubMed]
- Faure, S.; Georges, M.; McKey, J.; Sagnol, S.; de Santa Barbara, P. Expression Pattern of the Homeotic Gene Bapx1 during Early Chick Gastrointestinal Tract Development. Gene Expr. Patterns 2013, 13, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, C.; Murtaugh, L.C.; Chyung, J.C.; Lassar, A.; Roberts, D.J. Gizzard Formation and the Role of Bapx1. Dev. Biol. 2001, 231, 164–174. [Google Scholar] [CrossRef]
- Morgan, B.A.; Fekete, D.M. Manipulating Gene Expression with Replication-Competent Retroviruses. Methods Cell Biol. 1996, 51, 185–218. [Google Scholar] [CrossRef] [PubMed]
- Logan, M.; Tabin, C. Targeted Gene Misexpression in Chick Limb Buds Using Avian Replication-Competent Retroviruses. Methods 1998, 14, 407–420. [Google Scholar] [CrossRef]
- Cepko, C.L. Transduction of Genes Using Retroviral Vectors. In Current Protocols in Molecular Biology; Ausubel, F.M., Kingston, R.E., Moore, D.D., Seidman, J.G., Smith, J.A., Struhl, K., Eds.; John Wiley and Sons: New York, NY, USA, 1991; pp. 9:9.10.11–19.14.13. [Google Scholar]
- Harpavat, S.; Cepko, C.L. RCAS-RNAi: A loss-of-function method for the developing chick retina. BMC Dev. Biol. 2006, 6, 2. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody (Clone) | Host Species | Antigen Specificity | Dilution | Vendor (Catalog Number) |
|---|---|---|---|---|
| HNK-1 | mouse | IgM | 1:50 | Thermo Fisher, Waltham, MA, USA (MA5-11605) |
| Tuj1 (AA10) 1 | mouse | IgG2a | 1:100 | Santa Cruz, Dallas, TX, USA (sc-80016) |
| SMA (1A4) | mouse | IgG2a | 1:400 | Dako, Santa Clara, CA, USA (M0851) |
| p75NTR | rabbit (polyclonal) | IgG (H + L) | 1:300 | Promega, Madison, WI, USA (G3231) |
| pSMAD1/5/9 1 | rabbit (polyclonal) | IgG (H + L) | 1:200 | Cell Signaling, Danvers, MA, USA (#13820) |
| Bfabp | rabbit (polyclonal) | IgG (H + L) | 1:50 | kind gift of Dr. Thomas Müller |
| HuC/D (16A11) | mouse | IgG2b | 1:100 | Invitrogen, Waltham, MA, USA (A-21271) |
| N-cadherin (GC-4) | mouse | IgG1 | 1:200 | Sigma-Aldrich, St. Louis, MO, USA (C3865) |
| N-cadherin (6B3-c) | mouse | IgG1 | 1:5 | DSHB |
| SOX10 1 | mouse | IgG1 | 1:200 | Santa Cruz, Dallas, TX, USA (sc-365692) |
| RCAS gag protein (AMV-3C2) | mouse | IgG1 | 1:5 | DSHB |
| collagen III. (3B2) | mouse | IgG1 | supernatant | DSHB |
| collagen VI. (39) | mouse | IgG1 | supernatant | DSHB |
| fibronectin (B3/D6) | mouse | IgG2A | 1:3 | DSHB |
| Host | Target | Specificity | Vendor (Catalog Number) |
|---|---|---|---|
| horse | Anti-Mouse | IgG (H + L) | Vector Laboratories, Newark, CA, USA BA-2000 |
| goat | Anti-Rabbit | IgG (H + L) | Vector Laboratories BA-1000 |
| goat | Anti-Mouse | IgM (µ chain) | Vector Laboratories BA-2020 1 |
| Host | Target | Specificity | Excitation Wavelength | Vendor 1 (Catalog Number) |
|---|---|---|---|---|
| donkey | Anti-Mouse | IgG (H + L) | 488 nm | A21202 |
| donkey | Anti-Mouse | IgG (H + L) | 594 nm | A21203 |
| donkey | Anti-Mouse | IgG (H + L) | 647 nm | A31571 |
| donkey | Anti-Rabbit | IgG (H + L) | 488 nm | A21206 |
| goat | Anti-Mouse | IgG1 | 488 nm | A21121 |
| goat | Anti-Mouse | IgG1 | 594 nm | A21125 |
| goat | Anti-Mouse | IgG2a | 488 nm | A21131 |
| goat | Anti-Mouse | IgG2a | 594 nm | A21135 |
| goat | Anti-Mouse | IgG2b | 594 nm | A21145 |
| goat | Anti-Mouse | IgM | 594 nm | A21044 |
| goat | Anti-Mouse | IgM | 633 nm | A21046 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kovács, T.; Halasy, V.; Pethő, C.; Szőcs, E.; Soós, Á.; Dóra, D.; de Santa Barbara, P.; Faure, S.; Stavely, R.; Goldstein, A.M.; et al. Essential Role of BMP4 Signaling in the Avian Ceca in Colorectal Enteric Nervous System Development. Int. J. Mol. Sci. 2023, 24, 15664. https://doi.org/10.3390/ijms242115664
Kovács T, Halasy V, Pethő C, Szőcs E, Soós Á, Dóra D, de Santa Barbara P, Faure S, Stavely R, Goldstein AM, et al. Essential Role of BMP4 Signaling in the Avian Ceca in Colorectal Enteric Nervous System Development. International Journal of Molecular Sciences. 2023; 24(21):15664. https://doi.org/10.3390/ijms242115664
Chicago/Turabian StyleKovács, Tamás, Viktória Halasy, Csongor Pethő, Emőke Szőcs, Ádám Soós, Dávid Dóra, Pascal de Santa Barbara, Sandrine Faure, Rhian Stavely, Allan M. Goldstein, and et al. 2023. "Essential Role of BMP4 Signaling in the Avian Ceca in Colorectal Enteric Nervous System Development" International Journal of Molecular Sciences 24, no. 21: 15664. https://doi.org/10.3390/ijms242115664