Országos Pszichiátriai és Neurológiai Intézet 1021 Budapest, Hűvösvölgyi út 116. Az emberi élet csaknem egyharmada alvásból áll. Az alváskutatók ezen állapot számos neurofiziológiai és pszichológiai jellegzetességét tisztázták. Az alvás két fô formáját _ a REM és a NREM alvást _ különbözô neurofiziológiai folyamatok jellemzik. NREM alvás során a thalamo-corticalis interakció egy oszcilláló üzemmódra vált, melynek során az érzékszervi bemenetet ritmikus hiperpolarizációk gátolják. A REM alvást erôteljes neurális aktivitás jellemzi, ami a pons cholinerg struktúráiból ered és gátolja a thalamo-corticalis oszcillációt. Az agytörzsi monoaminerg és cholinerg sejtcsoportok kölcsönhatása a NREM és a REM alvás váltakozását szabályozza. Az alvás, mint egész a homeostaticus és a circadián folyamatok együttes kontrollja alatt áll. Jóllehet számos funkcionális hipotézis áll rendelkezésre, a NREM és a REM alvás pontos szerepe nagyrészt ismeretlen. Hipnagóg hallucinációk, gondolatszerű élmények és élénk álmodás jellemzik az elalvás, a NREM alvás és a REM alvás állapotait. Alvásnak nevezzük az élôlényeknek azt a majdnem az egész élôvilágban általános tulajdonságát, hogy bizonyos periodicitással, a környezettel való kapcsolatuk és motoros aktivitásuk nagymértékben csökken, speciesfüggô stereotip alváspozíciót vesznek fel (a denevér fejjel lefelé lógva, a lovak állva és az ember fekve) és mindez reverzibilis, a különbözô élôlényeknél különbözô idôtartam után ismét éber állapotnak adja át helyét. Az ember alvása durván az egész élettartam egyharmadát teszi ki, de az ébrenlét és alvás aránya életkorfüggôen jellegzetesen változik, csecsemô és kisgyermekkorban az alvás, idôskorban az ébrenlét javára. Az alvás és ébrenlét váltakozása az élôlények egyik legalapvetôbb biológiai ritmicitása.
Az NREM és REM alvás jellegzetességei és élettani hátterükEmberben, újszülött korban rövid alvás és ébrenlét periódusok váltják egymást a 24 órás napon belül számos alkalommal és az alvás tartama 17-18 óra. Késôbb az alvástartam fokozatosan csökken és 3-5 éves korra kialakul az alvás-ébrenlét ún. diurnális mintázata, vagyis az alvás egyre inkább egy nagyobb tömbben jelentkezik éjszaka, az alvás tartama 10-12 órára csökken, és nappal még mindig vannak alvás periódusok. Serdülôkorban alakul ki a felnôtt alvás-ébrenlét mintázat: 7-8 óra alvás egy tömbben éjszaka délutáni rövid alvás periódussal (szieszta) vagy anélkül. Idôskorban az éjszakai alvás tartama ismét csökken és a kisgyerekkori mintázatra emlékeztetve, nappal gyakori rövid felületes alvás periódusok jelentkeznek1. Az EEG, majd késôbb a polysomnographia bevezetésével egyre több objektív adat gyűlt össze az éjszakai alvás lefolyásáról és Aserinsky és Kleitman az 50-es évek elejérôl származó felfedezése óta ismert, hogy az alvásfolyamat két alvás típus, a NREM és a REM alvás, ciklikus váltakozásából épül fel (1. ábra).
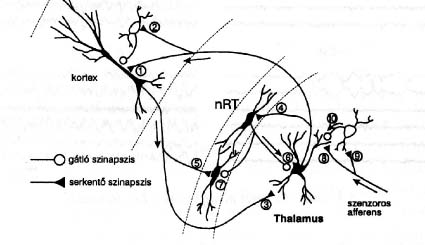
1. ábra Egy átlagos éjszakai alvás szakaszainak és fázisainak egymásutánja fiatal felnôttkorban. A NREM alvás alatti thalamo-corticalis interakció láncszemei. 1: corticalis piramissejt, 2: corticalis gátló interneuron, 3: corticalis piramissejt serkentô axonális összeköttetése a thalamikus relay sejttel, 4: a thalamikus relay sejt serkentô összeköttetése a thalamikus retikuláris magokkal, 5: a corticalis piramissejt serkentô összeköttetése a thalamikus reticularis magokkal, 6: a thalamikus reticularis magok gátló összeköttetése a thalamikus relay sejttel, 7: a thalamikus reticularis magok gátló összeköttetése egymás közt, 8-9: a thalamikus relay sejthez érkezô szenzoros afferens input, 10: gátló interneuron a szenzoros beáramlás és a thalamikus relay sejt között. NRT: nucleur reticularis thalami. A NREM alvást 90-120 percenként REM periódusok szakítják meg, melyek tartama estétôl reggelig nô és a REM-re jellegzetes állapotjellemzôk is egyre kifejezettebbek. Ugyanakkor viszont a NREM alvás „intenzitása” estétôl reggelig egyre csökken. Élettani körülmények között a REM alvás a felnôttek összalvásának mintegy 20-25 százaléka. A REM alvás tartama jellegzetesen életkorfüggô, újszülöttkorban 17-18 óra alvás mintegy 50%-át teszi ki és csak 2-3 éves korra csökken az alvás egy negyedére. Idôskorban tartama csak kevéssé csökken1-3. A NREM alvás vagy Lassú Hullámú Alvás (LHA) az agyi aktiváció és a somatikus működések jelentôs csökkenésével jár. Az EEG-ben lassú oscilláció és ún. alvási orsózás uralkodik, a szívfrekvencia és a légzésszám csökken, a vegetatív rendszer sympatikus uralmát parasympatikus túlsúly váltja fel, a testhômérséklet csökken. Egyes hormonok _ legpregnánsabban a növekedési hormon _ kiáramlása jellegzetesen lassú hullámú alvás függô. Az EEG változások a thalamikus nem-specifikus rendszer munkamódjának megváltozásával függnek össze. Ez a rendszer fázikus munkamódra vált, amelynek lényege az, hogy alternálóan izgalom és gátlás váltják egymást ciklikus módon mind a corticalis piramis, mind a thalamikus relé sejtekben. A gátló fázist a reticuláris thalamikus magrendszer fázikus GABA-erg gátló hatása biztosítja, amellyel mind a kéreg, mind a thalamus sejtjeire hat. A visszatérô gátlás meggátolja, hogy a thalamikus relé sejtek folyamatosan közvetíthessék a külvilág felôl a kéregbe áramló éberséget fenntartó ingerözönt (2. ábra).
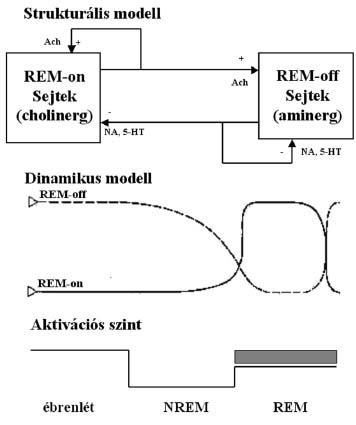
2. ábra A NREM alvás alatti thalamo-corticalis interakció láncszemei. 1: corticalis piramissejt, 2: corticalis gátló interneuron, 3: corticalis piramissejt serkentô axonális összeköttetése a thalamikus relay sejttel, 4: a thalamikus relay sejt serkentô összeköttetése a thalamikus retikuláris magokkal, 5: a corticalis piramissejt serkentô összeköttetése a thalamikus reticularis magokkal, 6: a thalamikus reticularis magok gátló összeköttetése a thalamikus relay sejttel, 7: a thalamikus reticularis magok gátló összeköttetése egymás közt, 8-9: a thalamikus relay sejthez érkezô szenzoros afferens input, 10: gátló interneuron a szenzoros beáramlás és a thalamikus relay sejt között. NRT: nucleur reticularis thalami. A thalamus működésváltozásának hatására csökken a frontális kéreg beidegzése és következésképpen metabolizmusa is. Ez az állapot egyúttal az agytörzsi cholinerg ébresztôrendszer működésének visszaszorulását is eredményezi és a kéreg beidegzésében mind a cholinerg mind az aminerg (szerotonin és noradrenalin) komponens jelentôs csökkenését hozza létre. A REM alvás, amely ciklikusan felváltja a NREM szakaszokat, teljesen más vonásokat mutat. Ebben az állapotban az agy éber állapotnál is nagyobb mértékben aktivált, ugyanakkor mind a sensoros beáramlás, mind a motoros rendszer gátlás alatt áll. Ezért valóban találó a „paradox alvás” megjelölés. A leglényegesebb vonások a következôk. Az EEG éber állapothoz, vagy még inkább igen felszínes 1-es stádiumhoz hasonlít. Az agyi átáramlás növekszik és nô az agy hômérséklete is. A kérgi kiváltott potenciálok komponens összetevôi az éber állapothoz hasonlóak. A vázizomzat, kivéve az oculomotor rendszert és a diafragmát, tónusvesztett. Ugyanakkor az arc és végtagok idôszakosan mioclónusosan ránganak és a szemgolyók sajátos nystagmusszerű, változó tartamú szünetekkel megszakított, rezgésben vannak. A szív és a légzés ritmusa gyors és különösen a szemmozgásos idôszakokban aritmiás szakaszokat tartalmaz. Férfiaknál penis, nôknél clitoris erekció van, függetlenül az álomtartalom sexualis jellegétôl. A REM-bôl ébresztett személyek mintegy 85%-a álmokról számol be. A thermoreguláció felfüggesztôdik, megszűnik az izzadás, egyben a NREM-re jellemzô gyakori electrodermalis tevékenység és poikilotherm állapot alakul ki.2,3 Agytörzsi átmetszéses és sértéses kísérletekkel kimutatták, hogy a REM állapotért felelôs struktúra a ponsban van és a kritikus terület a tegmentumban lateralisan a nucleus pontis oralist foglalja magába és a locus coeruleustól ventralisan helyezkedik el. Az itt elhelyezkedô sejtek REM alatt aktívan tüzelnek, míg LHA-ban és éber állapotban csendesek (REM-on neuronok). A REM kiváltható ezeknek a struktúráknak az acethylcholin agonista ingerlésével (pl. carbachol kristály implantálásával) is.2,4 Az alvás agyi szabályozásaAz alvásperiódus circadián szabályozásaNoha, amint fentebb láttuk, az alváson belül jellegzetes ultradian REM-NREM ciklusok vannak, az alvásfolyamat egy tömbben kezelhetô programként fogható fel. A kérdés, hogy mi indítja meg ill. idôzíti a nap egy bizonyos periódusában az alvásprogramot, régóta felmerült. Kérdéses volt, hogy az alvásciklus belsô, vagy külsô vezérlésű. Ennek eldöntésére kísérleti személyeket hat hónapig megfosztottak a külsô idôtagoló tényezôktôl, olyan speciális lakásokban éltek, ahol nem voltak ablakok, nem használhattak telefont, TV-ét, rádiót, nem volt órájuk és semmi kapcsolatuk nem volt a külvilággal. Ilyen körülmények között a kísérleti személyek alvásának circadián ciklicitása fennmaradt, de elalvási idejük mind késôbbre tolódott, a ciklusidô meghaladta a 24 órát és 25 órához közelített. Mindez amellett szólt, hogy az alvásperiódus ciklicitását külsô tényezôk ugyan valamelyest befolyásolják, de alapvetôen egy „belsô óra” szabályozása érvényesül. Feltételezhetô volt, hogy miután az alvás és ébrenlét a világosság- sötét váltakozásával szinkron jelenség az egész állatvilágban (függetlenül attól, hogy egyes állatok, mint éppen a patkány is, sötétben aktívak és világosban alszanak), a „circadián óra” összeköttetésben kell legyen a vizuális inputtal. Ocularisan injiciált tricialt aminosavak segítségével végül ez a retino-hypothalamikus pálya láthatóvá vált és ez vezetett el a suprachiasmalis mag circadián óra szerepének felfedezéséhez. Az alvásprogram idôzítését, vagyis a 24-órás napban való elhelyezkedését az elülsô hypothalamusban, a harmadik kamra elülsô recessusának kétoldalán, elhelyezkedô magrendszer a nucleus suprachiasmatis szabályozza. Ennek a magrendszernek a roncsolása patkányokban az alvásperiódusok jelentkezésének ritmusát teljesen felborította, viszont magzati suprachiasmalis magszövet implantálásával az alvásperiódusok korábbi ritmusa visszaállítható volt. Más oldalról viszont az alvás circadián ritmuson belüli idôzítését a melatonin szekréció szabályozza. A melatonin a corpus pinealeban szabadul fel. Szekréciója sötétség függô és az alvástól független. Sötétben szekretálódik mind az éjszaka alvó emberben, mind az éjszaka aktív állatokban és a világosság megszakítja termelôdését. A melatonin alvásidôzítést befolyásoló szerepe a suprachiasmalis magban érvényesül, amelyben melatonin receptorokat mutattak ki.2,4 Az alvás ultradián szabályozásaA REM-NREM ultradian ciklikus váltakozását bonyolult agytörzsi kapcsolatok szabályozzák. Feltételezhetô, hogy az agytörzsi cholinerg és adrenerg neuronok közötti reciprok interakció a ciklikus váltakozás leglényegesebb mozzanata. Az aminerg neuronok fokozatos elhallgatása felszabadítja a REM-ért felelôs cholinerg struktúrákat az általuk gyakorolt gátlás alól (3. ábra).
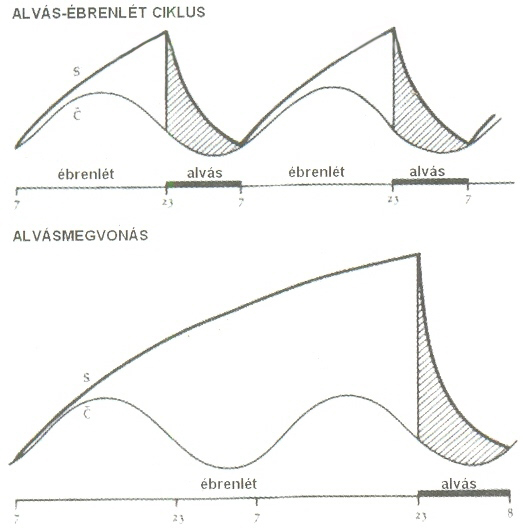
3. ábraStrukturális modell A cholinerg kisülések pedig egyrészt ingerületbe hozzák a thalamo-corticalis neuronokat, másrészt hiperpolarizálják a nucleus reticularis thalamit és ezzel felszabadítják az általa gyakorolt gátlás alól thalamo-corticalis neuronokat. Így megszakad a fázikus thalamo-corticalis gátlást szolgáltató kör és vele az LHA is, és érvényesülni kezd a REM. Az, hogy az aktuális REM periódus hogyan adja át a helyét ismét LHA-nak, egyelôre kevésbé kidolgozott. Az kétségtelen, hogy az aminerg beidegzés a kéregben legintenzívebb éber állapotban, legkisebb fokú REM-ben és a kettô között áll NREM-ben. Ugyanakkor a cholinerg beidegzés mind éber állapotban, mind REM-ben nagyfokú és lényegesen alacsonyabb szintű NREM-ben. Az ultradián szabályozás azonban nemcsak az alváson belül az LHA-t periódikusan megszakító REM szakaszok formájában érvényesül, hanem kimutatható az ébrenlétben tapasztalható ciklikus változások formájában is. Már Kleitman felvetette a 24 óra egészében érvényesülô mintegy 90 perces periódus idejű nyugalom-aktivitás ciklus („Basic Rest-Activity Cycle” _ BRAC) létezésének gondolatát. Késôbb egyes vizsgálók megerôsítô adatokat találtak egy, a REM-NREM váltakozáshoz hasonló nappali ritmus jelenlétére, az éberségi szint, egyes hormonszintek, vegetatív paraméterek, a motoros és kognitív aktivitás periodikus váltakozásában. Homeostatikus szabályozásA circadián és ultradian szabályozás mellett egyértelmű bizonyítékai vannak annak, hogy az alvásfolyamatot jelentôsen befolyásolja az elôzetesen ébren töltött idô tartama és milyensége is. Minél hosszabb idôt töltünk ébren, a rákövetkezô alvás annál több lassú hullámot tartalmaz, vagyis annál „mélyebb” lesz. Mintha az alvástól való különbözô mértékű megfosztottság valamilyen hiányt hozna létre, amit a következô alvásszakaszban be kell pótolnunk. Borbély és mtsai a 80-as évek elején kimutatták, hogy az ébren töltött idô függvényében az alvási EEG delta gazdagsága exponenciális görbe mentén növekszik. Alvásfosztott patkányokat vizsgálva azt találták, hogy alvás visszacsapást befolyásolta az is, hogy a 24 óra mely szakaszában fejezték be az állatok ébrentartását, és ebbôl arra következtettek, hogy a circadián nyugalom-aktivitás ciklicitás olyan kapuként szolgál, amely csak bizonyos periódusokban engedi, hogy a megnövekedett alvásigény érvényre jusson. Ebbôl fejlôdött ki az ún. „két folyamat” alvásregulációs elmélet, amely feltételezi, hogy az alvás és az ébredés idôzítését, valamint az alvás intenzitását két folyamat együttesen szabályozza. Az egyik folyamat homeostatikus természetű („S-process”), a másik az elalvás- és ébredés-készség circadián ritmicitásából („C-process”) ered. Az elalvás ott következik be, ahol az elalváskészség circadián sinusoid ingadozást mutató görbéjét az idôben exponenciálisan növekvô alvásigény görbéje metszi, és az ébredés ott, ahol az ébredéskészség az elôzetesen ébren töltött idô függvényében különbözô meredekségű exponenciálisan csökkenô delta teljesítménysűrűség görbéje metszi egymást.(4. ábra)
4. ábra Az alvásszabályozás Borbély-féle két-folyamat modellje Ezt az elképzelést matematikai modell formájában is kidolgozták, és számos alvásdeprivációval kapcsolatos jelenség jó egyezést mutatott a modell predikcióival. A Borbély féle „két-folyamat” modell az alvásciklusoknak csak a burkológörbéjét vette figyelembe, és még az egyes alvásfázisokban jelentkezô fluktuáció sem szerepelt tényezôként. Késôbb McCarley és Massaquoi (1992) revidiálták ezt a modellt, és ebbe már beépítették a külvilágból származó ébresztô inputot, és az alvás mikrostruktúrája egyre inkább szerepet kap a pathológiás alvások elemzésében is. Kiderült, hogy a pathológiás alvásjelenségek jelentôs része az arousal-függô mikrostrukturális fluktuációk kulcspontjain jelentkezik, és ezek az ingadozások predilekciós helyeket jelölnek ki és mintegy „kapuzzák” a pathológiás eseményeket2. Miért alszunk?Az alvás biológiai funkciója mindmáig nem tisztázott kellôképpen. Éppen ezért számos hipotézissel rendelkezünk, amelyek közül csupán a legmegalapozottabbakra szorítkozom. Nyilvánvalónak látszik hogy a két alvásforma, a REM és NREM, más-más biológiai funkciót szolgál. A NREM alvás leginkább idôtálló és egyben a laikus gondolkodással és mindennapi tapasztalattal is legjobban egybevágó hipotézise a „visszaállítási illetve visszanyerési” elmélet. Eszerint az alvás az ébrenlétben egyre inkább kimerülô illetve elhasználódó kémiai-fiziológiai folyamatok regenerációját segíti elô, valamilyen fajta pihenést szolgál. Ennek az elméletnek a legrobosztusabb alátámasztó adata az, hogy a növekedéssel fejlôdéssel járó életperiódusban, a gyermekkorban az alvás hosszabb illetve intenzívebb (értsd delta gazdagabb) és a növekedési hormon kiáramlása egyértelműen az alvás elején teljesített legmélyebb delta alvással esik egyidôbe, még akkor is, ha az alvás _ ébrenlét periódust felcseréljük. Ez az elképzelés egybevág a homeosztatikus szabályozásra vonatkozólag fentebb elmondottakkal is: eszerint az alvás folyamán, arányban az elôzetesen ébren töltött idôvel valami visszapótlódik, regenerálódik és minél hosszabb idôt töltünk elôzetesen ébren annál intenzívebb (deltagazdagabb) lesz az alvásfolyamat. Az elmélet predikciója szerint alvásban a fehérjeszintézis megnövekedését várhatnánk, azonban ezt nem sikerült egyértelműen kimutatni. Hasonlóképpen a fizikai igénybevétel következtében emelkedett katabolikus rátának együtt kellene járni az alvás idô vagy mélység fokozódásával, amit szintén nem tudtak a legtöbb erre vonatkozó vizsgálatban kimutatni. A másik közkeletű elképzelés szerint az alvás arra szolgál, hogy pihenô periódusban csökkentse a testhômérsékletet és a metabolikus rátát a melegvérű állatokban, ahol a hôháztartás belsô szabályozása nagy energiabefektetéssel jár. Valóban, az alvás során mintegy 10%-os metabolikus ráta csökkenés következik be, ez a kis energiamegtakarítás azonban nem valószínű, hogy magyarázhatja az egész alvásfolyamat phylogenetikai kialakulását. Kétségtelen azonban, hogy azonos elôagyi struktúrák és neurotransmitterek játszanak szerepet mind az alvás- mind a hômérséklet-szabályozásban, és hogy a hibernáció és az alvás között egy folyamatosság állapítható meg. Az alvás ökológiai hipotézise szerint a zsákmányállatok így kerülik el, hogy a ragadozók az éjszaka során felfigyeljenek rájuk, ás áldozattá váljanak. Ugyanakkor ebben az elképzelésben kevéssé érthetô, hogy miért kell ehhez olyan bonyolult folyamat, mint az alvás, hiszen a rejtôzés és a motoros aktivitás felfüggesztése is megtenné, és hogy ez a funkció miért maradt meg a phylogenezisben. Lehetséges, hogy a visszaállítási hipotézis kombinálható az ökológiaival, hiszen a regeneráció állapotában lévô szervezet kiszolgáltatottabb, kevésbé felkészült a ragadozók támadására és így az alvás kettôs _ de lehet hogy valójában többes _ célt szolgál. A REM alvás biológiai funkciója legalább olyan rejtélyes, mint a lassú hullámú alvásé. A funkcionális hipotézisek rendkívül széleskörűek. A REM alvás nagy aránya a fejlôdés korai szakaszában felvetette, hogy elôsegítheti az agy fejlôdését. Miután kimutatták, hogy a REM tartama növekszik kognitív terhelésre és egyes eredmények arra utaltak szerepe lehet a memória konszolidációban, a memória funkcióban és a kognitív elaborációban tulajdonítottak szerepet neki. Egy teljesen ellentétes elképzelés szerint, amelyet a Nobel-díjas Watson vetett fel, a REM fázis a felesleges memóriák törlését szolgálja. Felvetôdött, hogy a születéskor még nem kialakult fajspecifikus genetikai programozás folyik a REM fázisban. Egyik hipotézis sem nyert eddig egyértelmű bizonyítást. Annak ellenére, hogy a REM sajátos jelenségtartománya erôs kihívást jelent a kutatás számára, ez a kérdés nehezen megközelíthetô terület mindmáig és mintha az utóbbi idôben kissé meg is torpantak volna vizsgálatok, átütô eredmények nem születtek régóta.2,4 Az álom fiziológiája és pszichológiájaHypnagog hallucinációkAz alvásstádiumok electroencefalográfiás és polygráfiás jegyeinek ismeretében megvalósulhatott az álomélmények leíró és korrelatív pszichofiziológiai jellemzése. Megállapították, hogy az elalvást kísérô és követô néhány másodperces idôszakot élénk, vizuális, hallucinátoros élmények jellemzik, amelyek a megfelelô pillanatban történô, elôre megbeszélt ébresztések hiányában sohasem rögzülnek emléknyomokként. A hypnagog hallucinációk ritkán ábrázolnak az álomra jellemzô szcénikusan szervezett tartalmakat és még ritkábban történeteket. A narratív elemek hiánya a hypnagog hallucinációk és a tulajdonképpeni, REM alvásban keletkezô álmok közötti egyik legszembetűnôbb különbség. A hypnagog hallucinációk általában színek és formák bonyolult és gyorsan körvonalazódó keveredésébôl állnak, amelyeket nehéz valós tárgyaknak vagy személyeknek megfeleltetni. Tartalmuk igen változatos, ezért néha valós személyek, tárgyak és képi benyomások is fölbukkanhatnak bennük. Furcsamód szinte sohasem ábrázolják az alvó személyt, mint aktív résztvevôt. Ebben is jelentôsen különböznek a REM alvásra jellemzô álmoktól. A harmadik alapvetô különbség a hypnagog hallucinációk és az álmok között az, hogy az elôbbiek nagymértékben nélkülözik az érzelmi átélést, szemben a spontán ébredéskor vagy REM alvásból való ébresztéskor felidézett álmoktól, amelyet éppen érzelmi színezetük tesz emlékezetessé. Ezek a jelenségek a látókéreg elalváskori mindeddig ismeretlen eredetű aktivációjának jelei. Feltételezhetô, de távolról sem bizonyított, hogy az EEG regisztrátumokban elalvás környékén észlelhetô 80-200 msec idôtartamú, nagyfeszültségű (20-100 µV) theta sorozatok, az ún. pozitív occipitalis meredek tranziensek, ugyanannak a folyamatnak a mutatói. Ezek a hullámok nagyon hasonlítanak az ébrenlét során regisztrált lambda-hullámokhoz, amelyek erôsen megvilágított környezetben, vizuális explorációkor (képek nézegetése) jelentkeznek. Abból a megfigyelésbôl kiindulva, hogy a hypnagog hallucinációk általában az epizódikus memóriából (konkrét téri-idôi kontextusban rögzített emléknyomok rendszere) mozgósítanak szabad asszociációkat, mások a hypnagog hallucinációknak a memóriafolyamatokkal való kapcsolatára helyezik a hangsúlyt. Ezt a következtetést azonban kérdéses érvényességűvé tette az a megfigyelés, amely szerint a kétoldali hippocampus-károsodású, súlyos anterográd amnéziában szenvedô páciensek egy naponta több órán át gyakorolt videójáték képi szekvenciáit élték újra hypnagog hallucinációikban. Eközben sem a játékról, sem pedig a képek eredetérôl nem tudtak (epizódikus) emléknyomokat felidézni. Ez, a hippocampustól független, implicit memóriafolyamatoknak a hypnagog hallucinációkban való részvételét hangsúlyozó eredmény.4, 5 Mentális tevékenység a NREM alvásbanA mentális tevékenység a NREM alvás elôrehaladtával veszít az élénkségébôl. A 2, 3 és 4 alvásstádiumokban sokkal inkább fogalmi jellegű tartalmak töltik ki az alvó személy tudatát. A beszámolók 20%-a teljes mértékben nélkülözi a vizuális elemeket. Gyakran valamely konkrét dologra való emlékezést vagy annak gondolati felidézését tapasztalja meg az alvó. Mivel ezeket nem mindig éli meg a szó szoros értelmében vett álomként fontos, hogy kísérleti ébresztéskor a „Mit álmodott?” kérdés helyett a „Mi futott át az agyán, mielôtt fölébredt?” jellegű általánosabb kérdések hangozzanak el. Ez a tévedés idézte elô azt a ma is gyakran hangoztatott téves felfogást, amely szerint NREM alvásban hiányoznak a mentális tartalmak és a szubjektív átélések az alvó élményvilágából. Amennyiben képi elemek dúsítják a NREM alvást kísérô mentális életet, úgy azok általában kevésbé élesek és körvonalazottak, mint a hypnagog hallucinációkban és a REM-álmokban. Hangingerekkel azonban néha a 2 stádiumban szemmozgások válthatóak ki. Ilyenkor a vizuális élmények részvétele a REM alvásra jellemzôen fokozottabbá válik. A NREM alvást kísérô mentális aktivitás is nagyobb mértékben mozgósít epizódikus vagy önéletrajzi jellegű szabad asszociációkat. A hypnagog hallucinációktól eltérôen azonban ezeknek az élményeknek a tartalma is nagymértékben építkezik az alvó legutóbbi napjainak konkrét emlékeibôl. NREM alvás során, állatkísérletek tanúsága szerint az ébrenlét során szerzett tapasztalatoknak egyfajta idôben sűrített visszajátszása zajlik a hippocampusból a neocortex irányába. Lehetséges, hogy az alvó személy ennek a működésmódnak egy részleges szubjektív lecsapódását éli meg ilyenkor. Sajnos a hypnagog hallucinációktól eltérôen nem rendelkezünk adatokkal a hippocampus-lézióknak a NREM-mentációra gyakorolt hatásáról. A kérgi neuronok nagy része ritmikusan hyperpolarizálódik NREM alvásban és a thalamo-corticalis interakció is ritmikus gátló hatás alá kerül, miáltal az élénkebb és tartalmilag gazdagabb, ébrenlétre és REM alvásra jellemzô mentális élet ilyenkor nem válhat uralkodóvá. Ennek ellenére elôfordulhatnak álomszerű, bizarr asszociációk, hiszen a kéreg monoaminerg (noradrenerg és szerotoninerg) szabályozása az ébrenléthez képest alábbhagy.4 Az álmodás jellegzetességei és a REM alvással való kapcsolatukAz álmodás és a REM alvás kapcsolatának felismerése a modern alváskutatás egyik legjelentôsebb felfedezése. A spontán felidézett álmaink döntô többsége az alvás REM fázisaiban keletkezik. A REM alvásban keletkezô álomélmények hosszúak, szcénikusak, narratívak, érzelmi átélésekben gazdagok és általában kiemelkedôen bizarrak. Uralkodó érzékszervi modalitásuk a vizuális. Kisebb mértékben hallási (részben beszédhangra vonatkozó) és kinetikus-vestibularis reprezentációkat is tartalmaznak. Csak elenyészô mértékben építkeznek hôérzékelési, tapintásérzékelési, valamint szaglási és ízérzékelési szenzoros modalitásokból. Sohasem tartalmaznak explicit fájdalomérzékelést. Változatos érzelmi színezetükben a negatív emóciók dominálnak. Leggyakoribbak a szorongás, a félelem és a meglepôdés. Tartalmuk gazdag, változatos és hiperasszociatív jellegű, vagyis az egymással igen kevéssé kapcsolatos emlékek, képek, benyomások szokatlan, bizarr, sűrített kombinációját képezik. Kísérletileg kimutatták, hogy közvetlenül a REM alvásból való ébresztés után még fennáll egyfajta hiperasszociativitás a memóriafolyamatokban.6 Az álmok bizarrsága is elsôsorban a szokatlan tartalmak kombinációjából illetve a cselekmény dramaturgiájából fakad. Az álmok tartalma azonban mégis követ egy lazán szervezôdô következetességet abban az értelemben, hogy egyazon éjszaka ismétlôdô NREM mentációiban és álmaiban egyes tartalmak bizarr kontextusuk ellenére, hajlamosak ismételten felbukkanni, mintha ezek kiemelkedô fontossággal bírnának az álmodó számára. Az álomfolyamatok kognitív szervezôdésének legkiugróbb sajátossága minden bizonnyal a narrativitás vagy történetiség kizárólagosságának tendenciája. Szinte minden, amit álmodunk, történetek formájában ölt testet. Az álmodás neurobiológiai magyarázata érdekében az utóbbi évtizedekben jelentôs kísérleti és elméleti erôfeszítéseket tettek. Kétségtelen, hogy az álom tartalmi gazdagsága és vizuális élénksége összefüggésben áll az illetô REM fázis szemmozgás-sűrűségével. Úgy tűnik tehát, hogy az álmodást és a szemmozgásokat kiváltó agyi impulzusok legalábbis részben közösek: a REM alvást beindító hídi cholinerg struktúrák burst-szerű tüzelésével kapcsolatosak. Késôbb egy rendkívüli jelentôségű felfedezésben leírták a REM fázist kísérô egyik legkülönösebb neurofiziológiai folyamatot: az élettani részben már említett noradrenerg és szerotoninerg működéskimaradást. Részben ez jellemzi az adrenerg és hisztaminerg neuroncsoportokat is. Fontos kivételt képez a dopamin, amely nem szűnik meg felszabadulni REM alvás során. Ezeknek az aminerg sejtcsoportoknak kiemelkedô jelentôségük van a kognitív és az emocionális funkciók szabályozásában. A noradrenalin például a jel-zaj arány fokozásáról ismert neuromodulátorként viselkedô biogén amin. A szerotonin számos viselkedéses és pszichológiai funkció, többek között a limbikus rendszer gátlásának szolgálatában áll. Az említett biogén aminok tónusos befolyásának hiánya messzemenô következményekkel kell járjon az agyműködésben. A szerotonin-transzmisszió felfüggesztôdése például az LSD és más hasonló struktúrájú hallucinogének hatására ébren is bekövetkezhet, ami az ismert transzállapottal és vizuális hallucinációkkal jár együtt. Ez jól megmagyarázza az álomélet nehezen értelmezhetô sajátosságait. Két további tényezô járul hozzá ehhez a neurobiológiai magyarázathoz. Az egyik a relatív dopaminerg túlsúly, ami a hiányzó noradrenerg és szerotoninerg szabályozás mellett egyes feltételezések szerint egy átmeneti pszichózisszerű állapotot idéz elô, persze az alvás leple alatt. A fokozott dopaminerg tónust ugyanis a pszichotikus tünetekkel hozzák összefüggésbe a biológiai pszichiátriában, és a tünetek korrigálására leggyakrabban dopamin antagonistákat használnak. Ezzel szemben a dopamin agonisták mellékhatásként rémálmokat okozhatnak, nagyobb dózisban vagy túladagoláskor pedig pszichotikus tüneteket. A másik neurobiológiai alapú álomelmélet az agyi funkcionális képalkotó eljárások tapasztalatait és a neurológiai sérülések következtében elôálló álommódosulásokat használja fel. Kétségtelen, hogy az álmodás folyamata más agyi struktúrákon és azok másfajta kölcsönhatásán alapul, mint az éber szellemi élet. A híd, a limbikus-paralimbikus régiók és a magasabb rendű vizuális áreák kiemelkedô aktivitást mutatnak REM alvásban. A prefrontalis területek aktivitása az ébrenléthez képest alacsonyabb. Ez megfelel az emocionális tónus és a vizuális képzetek álombeli hangsúlyosságának, de nem árul el sokat a kölcsönhatás mikéntjérôl. A neuropszichológiai eredmények arra utalnak, hogy az álmodás az emlékek és érzelmek vizuális megjelenítéseként fogható föl, ami a kérgi aktiváció elülsô, basalis elôagyon keresztüli, útvonalán valósul meg. Azt jelenti mindez, hogy az absztrakt, neocorticalis régiókban tárolt emlékek, és az amygdala által mozgósított érzelmek visszavetülnek a vizuális információfeldolgozás parietalis és temporalis területeibe, ahol képekké alakulnak. Az electrofiziológiai eredmények azt sugallják, hogy a hippocampus kimeneti információtovábbítása REM alvásban nem jelentôs, ezért a közelmúlt emlékei csak nyomokban jelennek meg az álomban. Emiatt a felszínre kerülô régi emléknyomok nem rendelkeznek közelmúltbeli horgonypontokkal és kaotikusan szervezôdnek narratív élményekké. Az álmok hiperasszociatív jellege, egyes felsôbb szabályozási folyamatoktól való mentessége, valamint a régi emléknyomokból való építkezése különösen alkalmassá teszi ôket a pszichoterápiás munka során a páciens személyiségének megismerésére. Ezt általában az álomelemekre adott szabad asszociációk révén érik el. A szabad asszociációk ugyanis segítenek elhelyezni az álomelemeket a személy saját emlékezeti készletének egyedi struktúrájában. Elhelyezésük révén azután olyan kapcsolatokra és emlékezeti asszociációkra derülhet fény, ami a meglévô emlékek között áll fenn.4 Irodalomjegyzék:1. Halász P: Alvás és alvászavarok Medicina Budapest, 1982. 2. Halász P: Az alvás élettana in: Novák M.: Az alvás- és ébrenléti zavarok diagnosztikája és terápiája (pp 42-66) Okker, Budapest 2000. 3. Kukorelli T: Az ébrenlét és az alvás in: Ádám Gy. Fehér O.: Élettan biológusoknak (pp 928-43) Tankönyvkiadó, Budapest 1991. 4. Bódizs R: Alvás, álom, bioritmusok Medicina Budapest, 2000. 5. Stickgold R, Malia A, Maguire D, Roddenberry D, O’Connor M: Replaying the game: Hypnagogic images in normals and amnesics. Science 2000; 290. 350-3 6. Stickgold R, Scott L, Rittenhouse C, Hobson JA: Sleep-induced changes in associative memory. J Cogn Neurosci 1999 11. 182-193 7. McCarley, Hobson JA: Neural excitability modulation over the sleep cycle: A structural and mathematical model. Science 1975 189. 60 |